
Une hématie
Une hématie (érythrocyte) est une cellule du sang, un globule rouge, dont la fonction est de transporter l'oxygène vers les tissus respiratoires (poumons et branchies). Les hématies sont rouges car elles contiennent 280 millions de molécules d'hémoglobine.
Chez les mammifères, l'hématie s'appelle un érythrocyte, mais prend d'autres appellations chez d'autres animaux. Si les érythrocytes des mammifères sont anucléés (sans noyau) et sans organites, chez la plupart des autres vertébrés, le noyau et les organites subsistent dans les hématies.
Des hématies (érythrocytes) :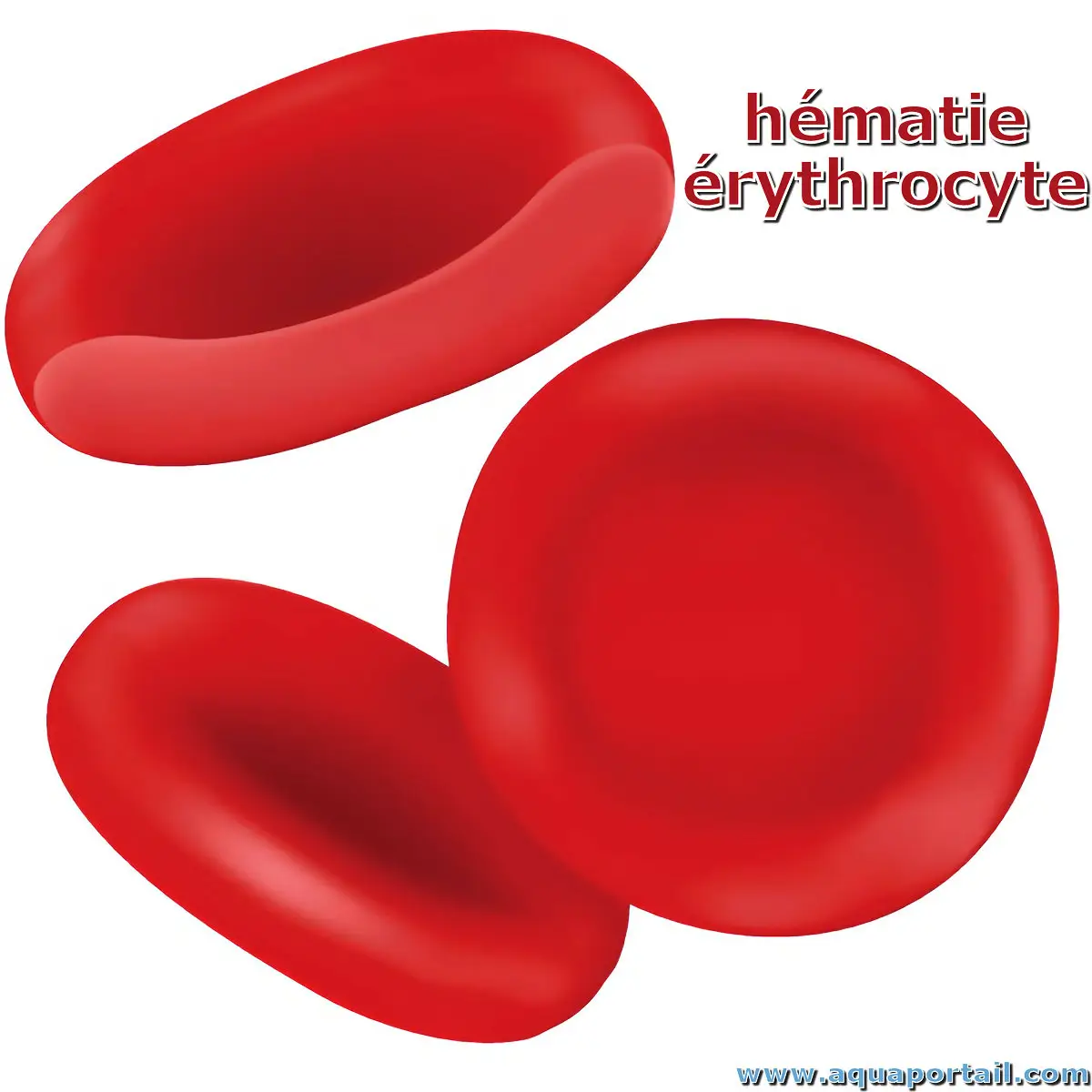
Les hématies sont un type de cellules sanguines fabriquées dans la moelle osseuse et présente dans le sang. Chez les mammifères, elles sont aussi appelées les érythrocytes.
Généralités
Les hématies, ou globules rouges, sont les cellules les plus nombreuses du sang. Elles ont une durée de vie courte, d'environ 4 mois. L'hémoglobine est l'un de ses composants principaux et sa fonction est le transport de l'oxygène O2 vers les différents tissus du corps. Une cellule du sang est appelée un hématocyte (dont les hématies font partie).
Les quantités d'hématies (érythrocytes) considérées comme normales chez les adultes sont (par μL -microlitres de sang-) : 4 à 5 millions/μL chez la femme, et 4,5 à 5,5 millions/μL chez l'homme.
Cette cellule sanguine, le globule rouge du sang, est sensible à la salinité du milieu à sa pression osmotique : elle peut être détruite par choc osmotique.
Les érythrocytes humains, ainsi que ceux des autres mammifères, n'ont ni noyau, ni mitochondries, de sorte qu'ils doivent obtenir leur énergie métabolique par la fermentation lactique. Les érythrocytes (globules rouges) complètent les leucocytes (globules blancs). Il y a environ 1000 fois plus d'érythrocytes que de leucocytes.
Le taux d'hématies dans le sang est un hématocrite, mesuré avec une prise de sang. L'excès d'hématies est appelé polycythémie ou polyglobulie (anomalie de la production des globules rouges lors de l'érythropoïèse) et le manque d'hématies est appelé anémie.
Érythrocyte
L'érythrocyte est un disque biconcave de 5 à 7,5 μm de diamètre, de 1 μm d'épaisseur et de 80 à 100 femtolitres de volume. La cellule a perdu son ARN résiduel et ses mitochondries, ainsi que certaines enzymes importantes; par conséquent, elle est incapable de synthétiser de nouvelles protéines ou de nouveaux lipides. Son cytoplasme contient principalement l'hémoglobine pigmentaire, qui lui donne sa couleur rouge caractéristique (qui peut être plus foncée selon son oxygénation) et est responsable du transport de l'oxygène (dioxygène).
Formes des diverses hématies :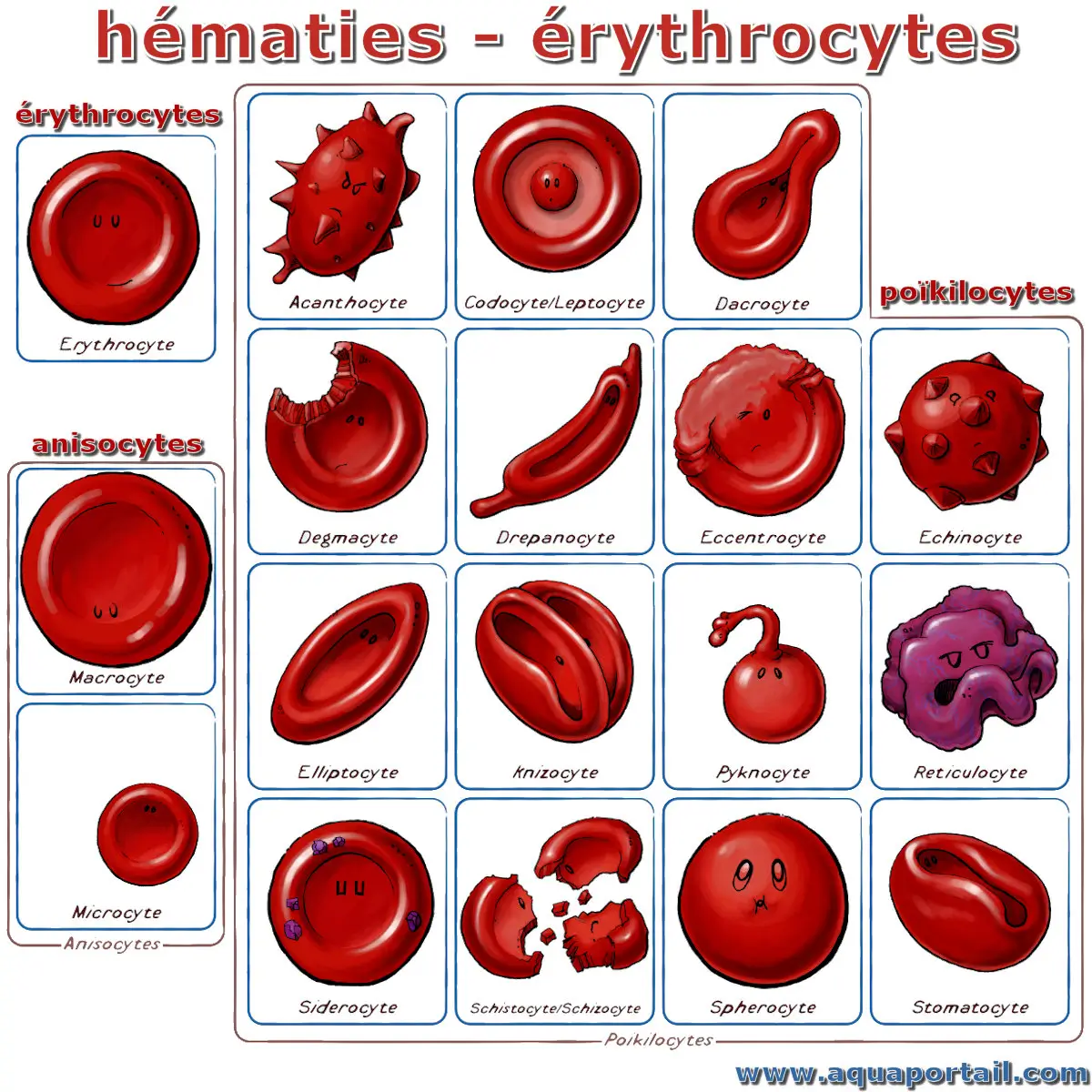
Les différents types d'hématies, dont les 3 principaux groupes : érythrocites, anisocytes et poïkilocytes.
Cependant, cette description est appliquée aux érythrocytes de mammifères, puisque chez le reste des vertébrés, à quelques exceptions près, les hématies n'ont pas la forme biconcave et tendent à être plus grandes. C'est parce que les hématies du reste des vertébrés ont encore un noyau.
Les érythrocytes sont dérivés des cellules souches compromises appelées hémocytoblastes. L'érythropoïétine, une hormone de croissance produite dans les tissus rénaux, stimule l'érythropoïèse (c'est-à-dire la formation d'érythrocytes) et est responsable du maintien d'une masse érythrocytaire dans un état constant. Les érythrocytes, comme les leucocytes, ont leur origine dans la moelle osseuse.
La concentration en érythrocytes varie selon le sexe, l'âge et la localisation géographique. Des concentrations plus élevées d'érythrocytes se trouvent dans les zones de haute altitude, chez les mâles et chez les nouveau-nés. Les diminutions en dessous de la plage de référence génèrent un état pathologique appelé anémie. Cette altération provoque une hypoxie tissulaire. L'augmentation de la concentration des érythrocytes (polycythémie, polyglobulie) est moins fréquente.
L'hémolyse est la destruction des érythrocytes âgés et elle intervient par les macrophages de la rate et du foie. Les éléments essentiels, globine et fer, sont conservés et réutilisés. La fraction hème de la molécule est catabolisée en bilirubine et en biliverdine et finalement excrétée dans le tractus intestinal et dans les urines. La rupture de l'érythrocyte au niveau intravasculaire libère l'hémoglobine directement dans le sang, où la molécule se dissocie en dimères α et β, qui se lient à la protéine de transport, l'haptoglobine. Ceci transporte les dimères au foie, où ils sont ensuite catabolisés en bilirubine et excrétés.
Chez les mammifères
Les érythrocytes des mammifères n'ont pas de noyau lorsqu'ils atteignent la maturité, c'est-à-dire qu'ils perdent leur noyau cellulaire et donc leur ADN; les amphibiens, les reptiles et les oiseaux ont des érythrocytes nucléé, avec un noyau. Les érythrocytes perdent également leurs mitochondries et utilisent le glucose pour produire de l'énergie grâce au processus de glycolyse suivi d'une fermentation lactique.
Des érythrocytes nucléés :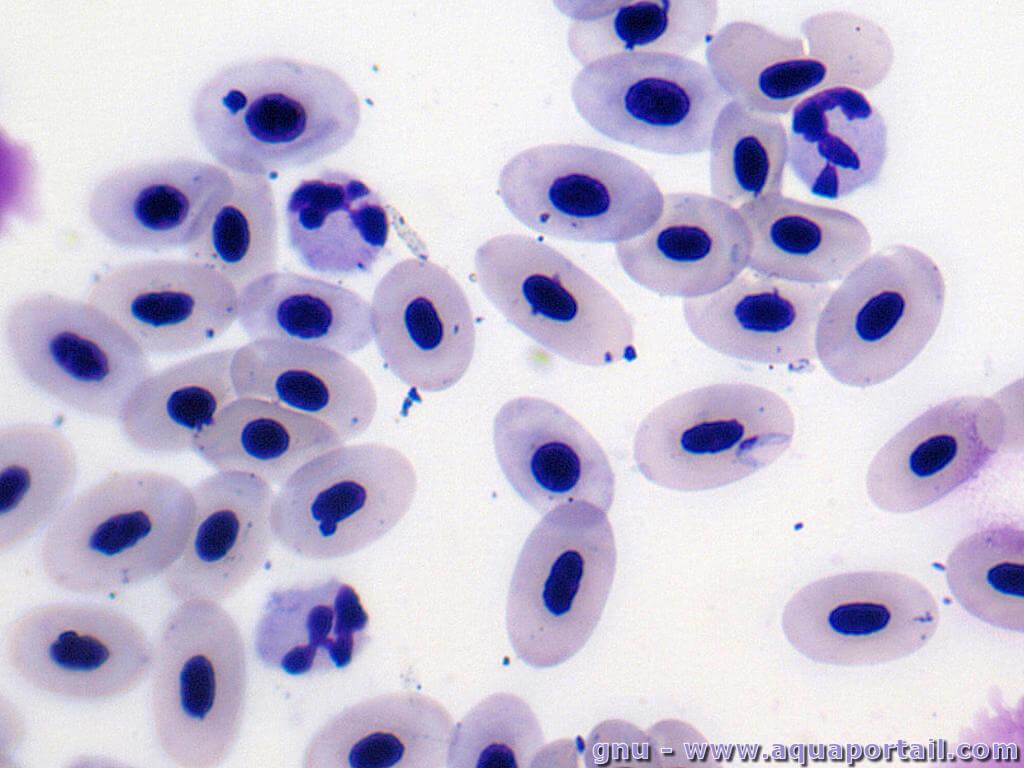
Les érythrocytes sont nucléés dans le sang des grenouilles.
Les érythrocytes sont produits en continu dans la moelle osseuse des os longs, bien que dans l'embryon, le foie soit le principal producteur d'érythrocytes. La rate agit comme un réservoir d'érythrocytes, mais sa fonction est quelque peu limitée chez l'homme.
Cependant, chez d'autres mammifères, tels que les chiens et les chevaux, la rate libère de grandes quantités d'érythrocytes lors des périodes de stress. Certains athlètes ont essayé d'exploiter cette fonction de la rate en essayant de libérer leurs réserves d'érythrocytes par des médicaments, mais cette pratique met en péril le système cardiovasculaire, car il n'est pas préparé à supporter un sang dont la viscosité est supérieure à la normale.
Chez les humains
Les érythrocytes humains ont une forme ovale, biconcave, aplatie, avec une dépression au centre. Cette conception est optimale pour l'échange d'oxygène avec l'environnement qui l'entoure, car elle leur donne la flexibilité de pouvoir traverser les capillaires, où ils libèrent la charge d'oxygène. Le diamètre d'un érythrocyte typique est de 6–8 μm. Les globules rouges contiennent de l'hémoglobine, qui est responsable du transport de l'oxygène et du dioxyde de carbone. En outre, c'est le pigment qui donne au sang la couleur rouge. La durée de vie d'une hématie-érythrocyte est d'environ 120 jours chez l'humain.
Vues d'hématies humaines :
Les hématies, érythrocytes ou globules rouges humains vus de la surface (a) sont ronds, mais forment des rouleaux vus de profil (b); ils ont un rendu sphérique (sphénocytes, c) par l'eau (hypotonique au sérum) et un rendu créné (échinocytes, d) par une solution saline (hypertonique). (c) et (d) n'interviennent normalement pas dans le corps.
Hémoglobine et hématies
L'hémoglobine est un pigment spécial qui donne aux hématies leur couleur rouge caractéristique. Sa molécule a du fer, et sa fonction est le transport de l'oxygène O2. Elle est présente chez tous les animaux, sauf chez certains groupes d'animaux inférieurs (avec des réticulocytes). Elle participe au processus par lequel le sang apporte les nutriments nécessaires aux cellules du corps et transporte ses déchets vers les organes excréteurs. Elle transporte également l'oxygène des poumons (ou des branchies chez les poissons et les nudibranches), où le sang le capte, vers les tissus du corps.
Lorsque l'hémoglobine se lie à l'oxygène pour le transporter vers les organes du corps, on parle d'oxyhémoglobine. Lorsque l'hémoglobine se lie au CO2 pour être éliminée par expiration, ce qui intervient dans les poumons, on parle de désoxyhémoglobine. Si l'hémoglobine se lie au monoxyde de carbone CO, il se forme un composé très stable appelé carboxyhémoglobine, qui a une très forte liaison avec le groupe hème de l'hémoglobine et empêche l'absorption d'oxygène, générant ainsi facilement anoxie qui conduit à la mort.
L'hémoglobine transporte également les déchets et le dioxyde de carbone dans les tissus. Moins de 2 % d'oxygène total et la plus grande partie du CO2 sont maintenus en solution dans le plasma sanguin. L'hémoglobine représente 35 % du poids de l'érythrocyte. Un composé apparenté, la myoglobine, agit comme un réservoir d'oxygène dans les cellules musculaires.
Maturation
Étant donné le besoin constant de reconstituer les érythrocytes, les cellules érythropoïétiques de la moelle osseuse sont parmi les cellules qui se développent et se reproduisent le plus rapidement dans le corps. Par conséquent, comme prévu, sa maturation et sa production sont très affectées en cas de carences nutritionnelles importantes.
Pour la maturation finale des érythrocytes, deux vitamines sont particulièrement nécessaires : la vitamine B12 et l'acide folique. Les deux sont essentiels pour la synthèse de l'ADN car tous deux, d'une manière différente, sont nécessaires à la formation de thymidine triphosphate, l'un des composants essentiels de l'ADN. Par conséquent, le manque de vitamine B12 ou d'acide folique provoque une diminution de la production d'ADN et, par conséquent, détermine un échec de la maturation et de la division nucléaires.
Une hématie (érythrocyte) en rouge :
L'hématie (érythrocyte ou globule rouge) apparaît à droite (en rouge) accompagnée d'une cellule de plaquette sanguine (thrombocyte, au milieu, jaune) et un lymphocyte T (à gauche, bleu) dans cette analyse d'image ultrastructurale tridimensionnelle du sang en utilisant un microscope électronique à balayage (MEB).
De plus, les cellules érythroblastiques de la moelle osseuse, en plus de ne pas proliférer rapidement, proviennent principalement d'érythrocytes plus grands que la normale appelés macrocytes, avec une membrane très mince, irrégulière et ovale, au lieu du disque biconcave habituel. Ces cellules malformées, après avoir pénétré dans le sang circulant, transportent normalement l'oxygène, mais en raison de leur fragilité, leur durée de vie est réduite de moitié à un tiers. Par conséquent, il est dit que le déficit en vitamine B12 ou en acide folique provoque une défaillance de la maturation érythropoïétique.
Il existe d'autres causes qui altèrent la maturation des érythrocytes, telles que la carence en fer et d'autres anomalies génétiques qui conduisent à la production d'hémoglobines anormales. Tous ces problèmes conduiront à des altérations des érythrocytes, par altération de la membrane, du cytosquelette ou autres.
Étapes du développement morphologique
Les étapes du développement morphologique de la cellule érythroïde comprennent (par ordre croissant de maturité) les étapes suivantes :
- Cellule souche pluripotente.
- Cellule souche multipotentielle.
- progénitrice ou cellule progénitrice ou les unités formant les colonies de la rate (voir les unités formant colonies) CFU-S.
- BFU-E (unité de formation des bourgeons érythrocytaires).
- CFU-E (unité de formation de colonies d'érythrocytes), qui formera ensuite les pro-érythroblastes.
- proérythroblaste : grande cellule de cytoplasme abondant, gros noyau avec chromatine épaisse, nucléoles pas très bien définis (20–25 microns).
- érythroblaste basophile : plus petit que le précédent (16–18 microns), cytoplasme basophile, chromatine grossière et grumeleuse, commence ici la formation d'hémoglobine.
- Érythroblaste polychromatophile : mesure 10–12 microns, le cytoplasme commence à acquérir une couleur rose en raison de la présence d'hémoglobine, ici la dernière phase mitotique pour la formation des globules rouges est présentée, il ne possède pas de nucléoles et le rapport noyau / cytoplasme est 4 : 1.
- Érythroblaste orthochromatique : mesure 8–10 microns, a la chromatine compacte et le noyau commence à disparaître.
- Réticulocytes : presque différenciés dans les érythrocytes matures. La présence de réticulocytes dans le sang périphérique) indique le bon fonctionnement de la moelle osseuse.
- Érythrocyte, enfin, quand il manque déjà de noyau et de mitochondries. Il a la capacité de transport (gaz, hormones, médicaments, etc.).
Au fur et à mesure que la cellule mûrit, la production d'hémoglobine augmente, entraînant une modification de la couleur du cytoplasme dans les échantillons de sang colorés avec le colorant de Wright, du bleu foncé au rouge et au rose rosé. Le noyau devient progressivement pycnotique et est expulsé de la cellule au stade orthochromatique.
La membrane érythrocytaire dans un complexe de protéine bilipidique, qui est important pour maintenir la déformabilité des cellules et la perméabilité sélective. À mesure que la cellule vieillit, la membrane devient rigide, perméable et l'érythrocyte est détruit dans la rate. La durée de vie moyenne de l'érythrocyte normal est de 100 à 120 jours.
Composition membranaire
La membrane érythrocytaire a plusieurs rôles qui aident à la régulation superficielle de la déformation, la flexibilité, l'adhésion à d'autres cellules et la reconnaissance immunologique. Ces fonctions dépendent fortement de leur composition, qui définit leurs propriétés. La membrane érythrocytaire est composée de trois couches : la glycocalyx à l'extérieur, riche en glucides; la bicouche lipidique qui contient plusieurs protéines transmembranaires en plus de leurs principaux constituants lipidiques; et le cytosquelette membranaire, un réseau structural de protéines situé sur la surface interne de la bicouche lipidique. La moitié de la masse de la membrane érythrocytaire chez l'homme et la plupart des mammifères sont des protéines, l'autre moitié sont des lipides, principalement des phospholipides et du cholestérol.
La membrane érythrocytaire est composée d'une bicouche lipidique, similaire à celle trouvée dans pratiquement toutes les cellules humaines. Cette bicouche lipidique est composée de cholestérol et de phospholipides, des lipides membranaires, en proportions égales en poids. La composition en lipides est importante car elle définit de nombreuses propriétés physiques telles que la perméabilité et la fluidité. De plus, l'activité de plusieurs protéines membranaires est régulée par l'interaction avec les lipides bicouches. Contrairement au cholestérol qui est réparti uniformément entre les monocouches interne et externe, les 5 phospholipides principaux sont disposés de manière asymétrique :
- Dans la monocouche externe :
- phosphatidylcholine;
- Sphingomyéline.
- Dans la monocouche interne :
La distribution asymétrique des phospholipides dans la bicouche est le résultat de la fonction de certaines protéines transporteuses de phospholipides à la fois dépendantes et indépendantes de l'énergie. Les flippases sont des protéines qui déplacent les phospholipides de la monocouche externe vers la monocouche interne, alors que les floppases font l'opération inverse, contre le gradient de concentration d'une manière dépendante de l'énergie. En outre, il y a les protéines de scramblase qui déplacent les phospholipides dans les deux directions en même temps, par leur concentration et leurs gradients indépendants de l'énergie. L'identité des protéines d'entretien de la membrane dans les érythrocytes est encore en discussion.
Le maintien de la distribution asymétrique des phospholipides dans la bicouche est critique pour l'intégrité et la fonctionnalité de la cellule pour plusieurs raisons :
- Les macrophages reconnaissent et phagocytent les érythrocytes qui ont exposé la phosphatidylsérine sur la surface externe. Par conséquent, le maintien de la phosphatidylsérine dans la monocouche interne est essentiel pour la survie de la cellule dans ses fréquentes rencontres avec les macrophages du système réticulo-endothélial, en particulier dans la rate.
- La destruction prématurée des érythrocytes thalassémiques et falsiformes a été liée à la désorganisation de l'asymétrie lipidique conduisant à l'exposition de la phosphatidylsérine dans la monocouche externe.
- Une exposition de phosphatidylsérine peut améliorer l'adhérence des érythrocytes aux cellules endothéliales vasculaires, empêchant efficacement le transit normal à travers la microvascularisation. Il est donc important de maintenir la phosphatidylsérine dans la monocouche interne de la bicouche pour assurer un flux sanguin normal dans la microcirculation.
- La phosphatidylsérine et le phosphatidylinositol-4,5-bisphosphate (PtdIns (4,5) P2) peuvent réguler la fonction mécanique de la membrane, en raison de leurs interactions avec des protéines cytosquelettiques telles que la spectrine et la protéine 4.1R. Des études récentes indiquent que la liaison de la spectrine à la phosphatidylsérine favorise la stabilité mécanique dans la membrane. PtdIns (4,5) P2 améliore la liaison de la protéine 4.1R à la glycophorine C mais diminue son interaction avec la bande protéique 3, et peut ainsi moduler la liaison de la bicouche au cytosquelette.
La présence de structures en radeaux spécialisées appelées radeaux lipidiques dans la membrane érythrocytaire a été décrite dans des études récentes. Ces structures riches en cholestérol et en sphingolipides sont associées à des protéines membranaires spécifiques, telles que la protéine G.
Métabolisme énergétique
Le métabolisme des érythrocytes est limité, en raison de l'absence de noyau, de mitochondries et d'autres organites subcellulaires. Bien que la liaison, le transport et la libération de l'oxygène et du dioxyde de carbone soient des processus passifs qui ne nécessitent pas d'énergie, il existe une variété de processus métaboliques dépendants de l'énergie qui sont essentiels à la viabilité de la cellule.
Les voies métaboliques les plus importantes pour les érythrocytes matures ont besoin de glucose comme substrat. Ces routes se réfèrent à:
Ces routes contribuent avec énergie en maintenant :
- potassium intracellulaire élevé, faible sodium intracellulaire et calcium intracellulaire très faible (pompe à cations);
- L'hémoglobine sous forme réduite;
- Des niveaux élevés de glutathion réduit;
- Intégrité et déformabilité de la membrane.
La glycolyse anaérobie (voie Embden-Meyerhof) fournit de l'ATP pour la régulation de la concentration intracellulaire de cations (Na+, K+, Ca2+, Mg2+) à travers des pompes à cations.
L'érythrocyte obtient de l'énergie sous forme d'ATP à partir du déroulement du glucose de cette manière. Les érythrocytes normaux n'ont pas de dépôts de glycogène, ils dépendent complètement du glucose environnemental pour la glycolyse. Le glucose entre dans la cellule grâce à une diffusion facilitée, un processus qui ne consomme pas d'énergie. Il est métabolisé en lactate, où il produit un gain net de deux moles d'ATP par mole de glucose.
Le cycle des pentoses fournit du nicotinamide-adénine dinucléotide phosphate et du glutathion réduit pour réduire les oxydants cellulaires. Environ 5 % du glucose cellulaire entre dans la voie oxydative des pentoses, un système auxiliaire pour produire des coenzymes réduites. Le glutathion réduit protège la cellule contre de nombreuses blessures causées par des agents oxydants permanents. Les oxydants dans la cellule oxydent les groupes sulfhydryle (-SH) de l'hémoglobine, à moins que les oxydants ne soient réduits par le glutathion réduit. C'est pourquoi le rôle de cette voie est crucial dans l'érythrocyte.
La voie de l'hémoglobine réductase protège l'hémoglobine de l'oxydation via le NADH et la méthémoglobine réductase. C'est une voie alternative à la voie d'Embden-Meyerhof, essentielle pour maintenir le fer hémique dans l'état Fe2+ réduit. L'hémoglobine avec le fer ferrique, Fe3+, est connue sous le nom de méthémoglobine. Cette forme d'hémoglobine ne peut pas être combinée avec l'oxygène. La méthémoglobine réductase, en association avec le NADH produit par la voie d'Embden-Meyerhof, protège le fer hémique de l'oxydation. Sans ce système, 2 % de la méthémoglobine formée tous les jours augmenterait, au fil du temps, à 20–40 %, limitant sévèrement la capacité de transport d'oxygène dans le sang. Les médicaments oxydants peuvent interférer avec la méthémoglobine réductase et produire des valeurs de méthémoglobine encore plus élevées. Cela provoque une cyanose.
Le cycle Rapoport-Luebering fait partie de la voie Embden-Meyerhof et vise à prévenir la formation de 3-phosphoglycérate et d'ATP. Le BPG (2,3-bisphosphoglycérate) est présent dans l'érythrocyte à une concentration de 1 mole BPG/mole d'hémoglobine et se lie fortement à la désoxyhémoglobine, de sorte que l'hémoglobine reste à l'état désoxygéné et facilite la libération d'oxygène. L'augmentation de la concentration en diphosphoglycérate facilite la libération d'oxygène dans les tissus en diminuant l'affinité de l'hémoglobine pour l'oxygène. De cette manière, l'érythrocyte possède un mécanisme interne pour réguler l'apport d'oxygène aux tissus.
En rapport avec "hématie"
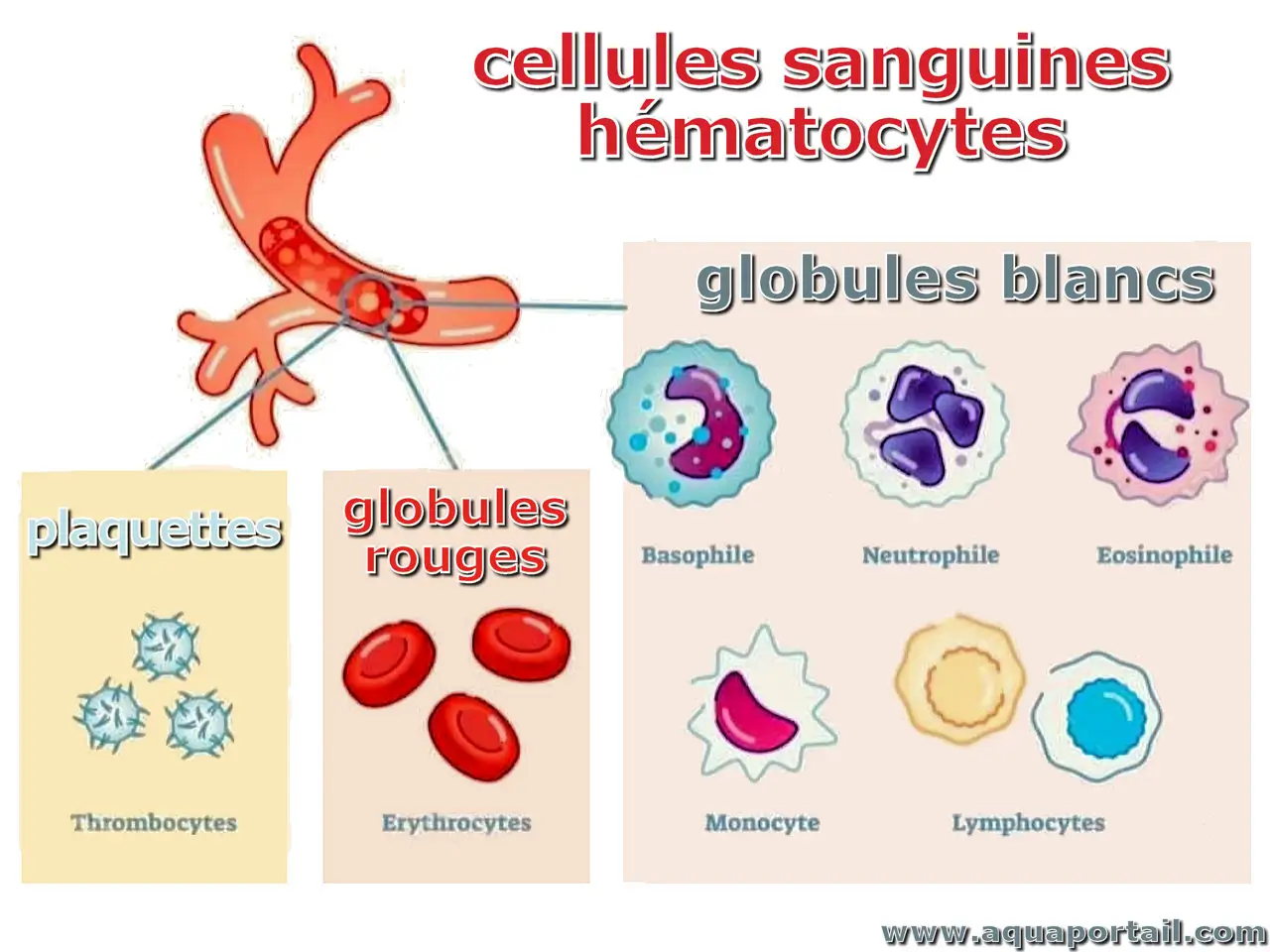
Une cellule sanguine (hématocyte) est l'un des trois organites du sang (plaquettes sanguines, globules blancs, globules rouges) produits par hématopoïèse.

Un globule rouge (érythrocyte, hématie) est le principal élément du sang, sous la forme d'un globule sanguin.
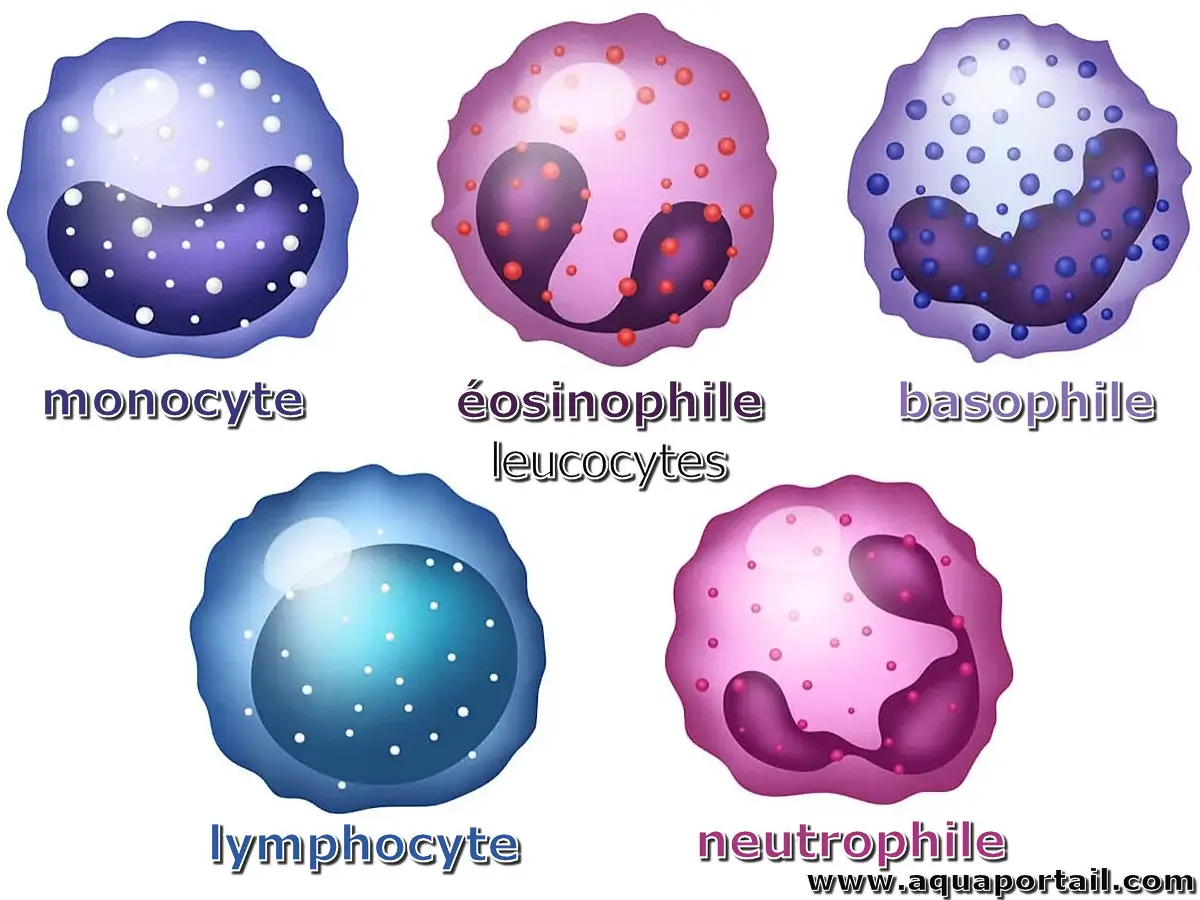
Les leucocytes regroupent un ensemble de cellules qui régissent l'immunité innée et adaptative.
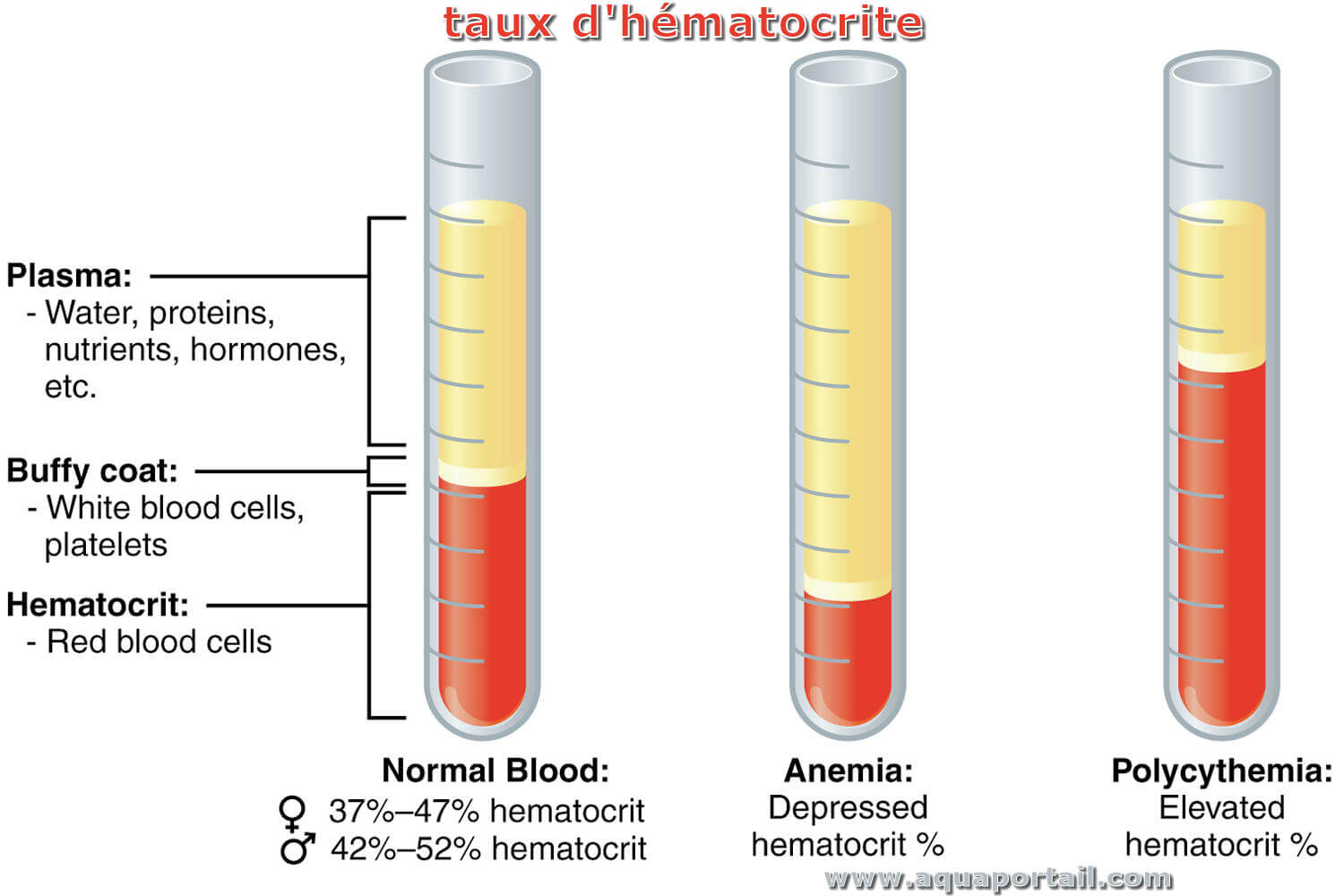
Un hématocrite est le rapport du volume des globules sanguins (hématies) sur le volume sanguin total.