
L'acide abscissique
L'acide abscissique est une hormone régulatrice de croissance naturelle impliquée dans de nombreux processus physiologiques avec la dormance des semences et des bourgeons, l'abscission des fleurs, les feuilles et fruits, le remplissage des semences, etc. L'abréviation ABA est très usitée pour cette phytohormone qui joue un rôle important dans la physiologie de la plante. Ses synonymes anciens sont les termes abscissine et dormine.
L'acide abscissique avec les stomates :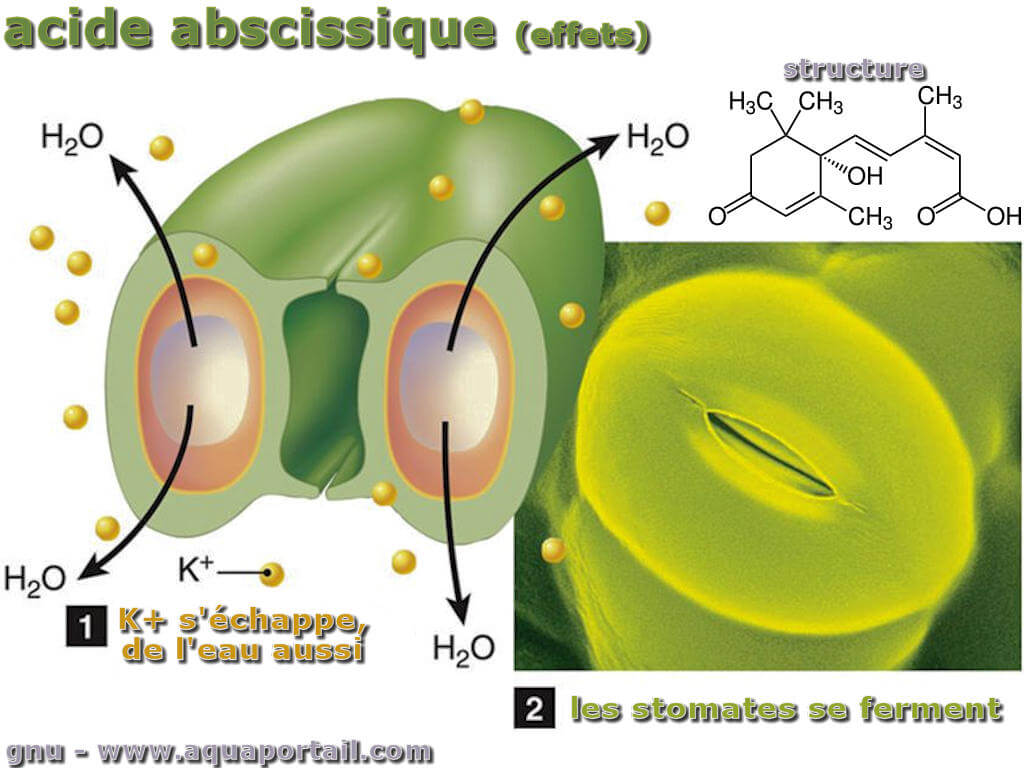
L'acide abscissique apporte une réponse à divers stress notamment au stress osmotique (stress dû à la sécheresse, à la chaleur ou au sel).
Généralités
L'acide abscissique est un sesquiterpène (à 15 °C) synthétisé par les végétaux supérieurs. Il ralentit la croissance, provoque la fermeture des stomates, favorise la maturation et la dormance des graines et des bourgeons, accélère la sénescence. Il accélère la chute des feuilles et des fruits mais ne la déclenche pas. Sa synthèse augmente en période de stress : sécheresse, anoxie des racines, forte concentration en sels. Cet acide participe aux processus de développement et de croissance ainsi qu'à la réponse adaptative aux stress biotiques et abiotiques.
Comme hormone végétale, l'acide abscissique inhibe généralement la croissance et favorise la dormance, aide la plante à résister aux conditions défavorables. Pour cette raison, elle est appelée aussi hormone de détresse. Elle serait responsable de l'entrée en dormance des graines et bourgeons à l'approche de la mauvaise saison, et de la fermeture des stomates en cas de déficit hydrique de la plante.
D'autres phytohormones que l'ABA sont : l'auxine (eau de saule), la gibbérelline, l'éthylène, les cytokinines et les brassicostéroïdes.
L'ABA est présent chez les plantes supérieures, les mousses, les algues, les champignons et les cyanobactéries, mais pas chez les autres bactéries, les archées ni les hépatiques.
L'acide abscissique a été découvert au début des années 1960, lorsque son implication dans le contrôle de la dormance des graines et de l'abscission d'organes a été découverte. De nos jours, on sait que l'éthylène est en réalité l'hormone qui intervient principalement dans l'abscission d'organes et que l'abscission d'organes induite par l'ABA observée dans les fruits du coton est due. à la capacité de l'ABA à induire la synthèse d'éthylène.
Schéma expliqué
L'acide abscissique apporte une réponse à divers stress. Un stress osmotique (stress dû à la sécheresse, stress dû à la chaleur, stress dû au sel) : limitation de la transpiration (l'ABA induit la fermeture des stomates via des flux ioniques, réduisant ainsi l'évaporation de l'eau) et limitant la photosynthèse. La phytohormone ABA induit l'ouverture de canaux anioniques au niveau moléculaire, ce qui permet aux anions de s'échapper de la cellule en fermeture. En conséquence, le potentiel de membrane devient plus positif (dépolarisation), ce qui ouvre à son tour les canaux de distribution de potassium et les ions K+ s'écoulent de la cellule vers l'apoplaste. L'eau de la cellule s'écoule dans l'apoplaste, la turgescence dans les cellules de garde diminue et les stomates se ferment. En cas de stress sec (stress hydrique), la concentration en ABA induite par le stress est en corrélation linéaire avec la conductivité hydrique des feuilles lorsque l'efficacité d'utilisation de l'eau augmente.
Structure chimique
L'ABA est un sesquiterpénoïde de 15 atomes de carbone C15H20O4. C'est un carbone anomère qui donne lieu à l'existence de deux énantiomères (S o + et R o -). La forme S est celle qui est présente dans la nature. Dans de nombreux processus physiologiques, la forme R est faiblement active, alors que dans d'autres, la forme R non naturelle présente une activité comparable à la forme S. Son nom systématique est (S) -5- (Acide 1-hydroxy-2,6,6-triméthyl-4-oxo-1-cyclohexyl) -3-méthyl-cis, trans-penta-2,4-diénoïque.
La chaîne latérale de la molécule ABA contient deux doubles liaisons conjuguées à l'extrémité carboxy-terminale. Le groupe le plus éloigné du groupe fonctionnel est en conformation trans, tandis que le groupe le plus éloigné est en configuration cis. L'orientation du groupe carboxyle de C2 conduit donc à l'existence d'une isomérie cis-trans. Presque tout l'ABA présent dans la nature est la forme cis et, par convention, le nom d'ABA est à propos de cet isomère.
Le groupe carboxylique donne également le caractère acide de la molécule ABA. Il s'agit donc d'un acide faible avec un pKa de 4,7. À cette valeur de pH, la forme protonée ABA-H est à la même concentration que la forme ionique chargée. La forme protonée peut être transportée à travers les membranes biologiques, mais pas la forme ionique chargée. Cependant, à un pH physiologique de 7,7, la forme anionique chargée est prédominante, elle existe à une concentration plus de 1 000 fois supérieure à la forme protonée. Étant donné que cette forme d'ABA ne peut pas pénétrer dans les membranes, il est nécessaire de disposer d'un système de transport permettant à l'ABA de transloquer entre les cellules.
Contrairement à la plupart des autres phytohormones, l'acide abscissique est une substance unique et non un groupe de substances.
Biosynthèse
L'ABA est synthétisée principalement dans les plastides des tissus vasculaires. Les dernières étapes de sa synthèse ont lieu dans le cytosol de la cellule. L'ABA est essentiellement dérivé d'un composé 5C appelé pyrophosphate d'isopentényle (ou pyrophosphate d'isopentényle), qui provient de la condensation d'une molécule de pyruvate et d'une autre du glycéraldéhyde-3-phosphate présent dans les plastides. L'isopentényl pyrophosphate se poursuit par la voie 2M-méthyl-D-érythritol-4-phosphate (MEP) ou par la voie indépendante du mévalonate jusqu'à la formation d'un composé intermédiaire appelé z40x - zéaxanthine.
La zéaxanthine est scindée en unités plus petites pour donner de la violaxanthine avec de l'anthoxanthine en tant que composé intermédiaire. Ces étapes sont catalysées par une zéaxanthine époxydase. Ensuite, des isomères de ce composé, tels que la cis-violaxanthine, ainsi qu'un nouveau composé, la néoxanthine et son isomère, sont formés. Vraisemblablement, ces étapes impliquent l'action d'une néoxanthine synthase et d'une isomérase. Les composés 9'-cis-néoxanthine et 9-cis-violaxanthine (9-cis-époxycaroténoïdes), par l'action d'une dioxygénase 9-cis-époxycaroténoïde (NCED), donnent naissance à la xanthonine, de 15 atomes de carbone, qui quitte les plastides vers le cytosol de la cellule. La xanthonine est ensuite convertie en un aldéhyde d'ABA par l'action d'une alcool déshydrogénase à chaîne courte (ABA2 chez Arabidopsis).
Enfin, cet aldéhyde d'ABA subit un processus d'oxydation par une aldéhyde oxydase d'abscis (AAO3) pour donner naissance à la molécule d'acide abscissique.
Catabolisme de l'acide abscissique
Les niveaux d'ABA sont régulés par un équilibre continu entre les formes actives et inactives, ce qui est très important dans la réponse de la plante au stress. Cet équilibre est atteint non seulement grâce à la synthèse et au catabolisme de l'ABA, mais également aux processus de conjugaison et de déconjugaison. Cependant, la synthèse et le catabolisme sont les principaux mécanismes impliqués dans la régulation des niveaux d'ABA dans la plante.
Le catabolisme de l'ABA inclut les processus de conjugaison qui inactivent la molécule d'ABA. Les processus du catabolisme comprennent deux voies principales :
- Voie oxydante : ils génèrent de l'acide phaséique et de l'acide dihydroxyphase. Ces composés n'ont pratiquement aucune activité. Elles sont produites par hydroxylation de la molécule ABA en position C8 'par des monoxigénases de type P-450 de la famille CYP707A (ABA 8'hydroxylases) pour générer un intermédiaire instable, 8'-OH-ABA, qui est isomérisé. à l'acide phaséique. Bien que l'hydroxylation en 8 'soit la voie catabolique principale, l'ABA peut également être hydroxylé aux positions 7'et 9 '. 12 Dans la phase de réhydratation après un stress hydrique, il a été constaté que, si les niveaux d'ABA diminuent, ceux d'acide phaséique augmentent.
- Conjugaison : l'ABA ou ses métabolites peuvent être inactivés par conjugaison avec une autre molécule. Le conjugué le plus courant est l'ester glycosylique ABA (ABA-GE), qui est formé par une réaction d'estérification effectuée par une glycosyltransférase. Contrairement à la voie oxydative, l'inactivation de l'ABA par conjugaison avec du glucose est un processus réversible. L'hydrolyse de l'ABA-GE est réalisée par une β-glucosidase et entraîne la libération de l'ABA. De plus, la forme ABA-GE n'est pas seulement une forme de stockage ABA mais également un moyen de transport. ABA-GE s'accumule dans les vacuoles et l'apoplaste, mais est transporté vers le réticulum endoplasmique en réponse à la déshydratation.
Fonctions
L'ABA participe activement à de multiples processus physiologiques de la plante, tels que la maturation de l'embryon, la dormance des graines, la croissance végétative et les processus liés à la tolérance au stress, tant biotique qu'abiotique. La grande importance de l'ABA dans la plante est claire lorsque nous observons le rôle que cette phytohormone exerce sur l'expression des gènes, ayant observé dans une expérience de microréseau (puces à ADN) que l'ABA est capable de modifier le profil d'expression de près de 10 %. des gènes d'Arabidopsis. En réalité, nombre de ces gènes sont probablement régulés par l'ABA.
L'acide abscissique est généralement antagoniste aux phytohormones stimulant la croissance et constitue donc un inhibiteur de croissance naturel. Avec les auxines, les gibbérellines et les cytokinines qui favorisent la croissance, l'acide abscissique régule le vieillissement, la carie des feuilles, la floraison, la maturation des fruits, le repos des graines et des bourgeons, la transpiration stomatique et d'autres processus de développement des plantes.
Croissance, développement et germination de la plante
L'ABA est nécessaire pour contrôler "finement" la croissance et le développement dans des conditions normales et sans stress. Cela est particulièrement important dans les processus qui interviennent dans la graine, tels que la dormance et la maturation de l'embryon. La dormance de la graine permet de retarder sa germination jusqu'à ce que les conditions du milieu environnant soient optimales pour la survie de la génération suivante.
La teneur en ABA des graines est généralement faible pendant les premiers stades de l'embryogenèse et atteint son maximum vers le milieu du processus pour redescendre plus tard. Pendant la maturation, il y a deux pics (maximum) d'accumulation d'ABA, l'un d'origine maternelle à 9–10 DAF (jours après la floraison, plusieurs jours après la floraison), avant le stade de maturation et l'autre d'origine embryonnaire à le 15–16 DAF. Le pic d'origine maternelle aide à prévenir la germination prématurée de l'embryon, tandis que le pic d'origine embryonnaire induit la dormance et la tolérance à la dessiccation, régulant ainsi la synthèse des protéines impliquées dans ces processus. Cette augmentation des niveaux d'ABA est déterminée par les processus qui influencent l'homéostasie (biosynthèse, transport et catabolisme).
Pendant la phase d'embryogenèse, la graine subit un processus de séchage qui implique la perte de jusqu'à 90 % de la teneur en eau de la graine, ce qui conduit à un état de dormance en dormance. L'embryon est capable de tolérer cette situation grâce à l'accumulation de solutés compatibles et de protéines déhydrinas et LEA (embryogenèse tardive abondante). Ces protéines sont de petites molécules hydrophiles qui s'accumulent tardivement au cours du processus d'embryogenèse. Elles retiennent l'eau et protègent les membranes et les autres protéines contre les dommages pouvant être causés par la faible disponibilité en eau. L'ABA participe directement à ces processus, car elle est capable d'induire la synthèse des protéines LEA et, par conséquent, d'apporter à la graine une tolérance à la dessiccation.
La diminution de la quantité d'ABA dans la graine par son catabolisme libère la graine de cette dormance et permet sa germination. Cependant, comme dans la plupart des processus physiologiques chez les plantes, l'ABA n'est pas la seule phytohormone qui participe à ce processus, mais d'autres comme les gibbérellines, l'éthylène et les brassinostéroïdes.
L'ABA a également un effet inhibiteur sur la croissance des racines latérales sous stress salin.
Stress abiotique
Le rôle de l'ABA dans les mécanismes de tolérance au stress abiotique de la plante a fait l'objet de nombreuses études car il limite considérablement la productivité des cultures d'importance agronomique. Le stress abiotique ayant la plus forte incidence est le déficit en eau, une condition environnementale relativement fréquente dans certaines régions de la planète. La sécheresse n'est pas le seul stress abiotique qui cause un déficit en eau pour la plante, mais d'autres types de stress abiotiques tels que la salinité, le gel ou les températures élevées entraînent également un déficit en eau.
Les conditions de salinité élevée entraînent un déficit en eau car le potentiel hydrique du sol (l'eau disponible pour la plante) diminue, car le potentiel osmotique d'une solution saline est inférieur à celui de l'eau, ce qui rend difficile l'absorption des racines par l'eau. Une salinité élevée augmente les niveaux d'ABA dans la plante.
Les températures élevées augmentent les pertes en eau par la transpiration, provoquant un stress hydrique dans la plante.
La congélation provoque un stress osmotique car la formation de cristaux de glace dans les espaces extracellulaires réduit le potentiel hydrique, ce qui entraîne la libération d'eau par les cellules, ce qui provoque une plasmolyse et finalement la mort cellulaire.
L'ABA aide à lutter contre le stress hydrique sur plusieurs fronts. Quand il y a un stress hydrique, les niveaux d'ABA dans l'usine augmentent. L'ABA est capable d'induire la fermeture des stomates afin de minimiser les pertes d'eau par transpiration. La fermeture de la stomie est réalisée par des modifications du potentiel de la membrane et du flux ionique, ce qui entraîne des modifications de la turgescence des cellules occlusives et par conséquent de leur fermeture. 3 Cependant, la fermeture des stomates présente des aspects négatifs tels que des échanges gazeux plus faibles, affectant négativement le taux de photosynthèse, entraînant une diminution de la disponibilité biologique des photoassimilats.
L'ABA induit également la synthèse de solutés compatibles (osmolytes, généralement de petites molécules, solubles, non chargées et sans danger à des concentrations élevées, n'affectant donc pas la fonction cellulaire que la proline, la glycine-bétaïne et le tréhalose. Ces solutés réduisent le potentiel hydrique des cellules, permettant ainsi de capter davantage d'eau environnante ou de conserver ce qu'elles possèdent. Les solutés compatibles augmentent également la stabilité et l'intégrité des membranes et des protéines, prévenant ainsi les dommages aux cellules.
Si les conditions de stress hydrique se prolongent dans le temps, certaines protéines LEA s'accumulent, empêchant ainsi leur agrégation ou facilitant leur repliement. Certaines de ces protéines LEA sont également induites par l'ABA.
Stress biotique
Le rôle de l'ABA dans la résistance aux maladies est complexe à établir car il varie en fonction du tissu, du stade de développement de la plante et du type d'interaction. Malgré cela, il est possible de dégager un schéma général suggérant un rôle stimulant de l'ABA dans la défense de la plante au début de l'infection, mais principalement négatif au cours des derniers stades.
Le rôle de l'ABA pendant les stades pré-invasifs est donné par sa fonction dans la fermeture des stomates. En raison de sa position d'interphase entre l'intérieur de la plante (tissus de la plante) et l'extérieur (environnement), les stomates sont une passerelle pour la colonisation endophyte par certains phytopathogènes où la fermeture de stomie dépendante de l'ABA il est susceptible de fonctionner comme une barrière de défense pré-invasive contre certains agents pathogènes virulents.
L'ABA exerce également des effets aux derniers stades de l'infection, soit en supprimant la résistance, soit en améliorant la sensibilité de la plante. Cela est dû au rôle antagoniste de l'ABA vis-à-vis des principales hormones impliquées dans l'interaction plante-pathogène, telles que jasmonique, salicylique et éthylène.
Propriétés
L'acide abscissique forme des cristaux incolores facilement solubles dans les solvants organiques. La solubilité dans l'eau dépend du pH : dans les solutions acides et neutres, l'acide abscissique se dissout mal, dans les puits alcalins. L'acide abscissique est très sensible à la lumière. Lors de l'exposition à la lumière, un réarrangement photochimique en isomère trans physiologiquement inactif survient.
Transport de l'ABA à travers la membrane
La capacité de l'ABA à diffuser à travers la membrane réside dans sa nature acide faible. L'ABA a un pKa de 4,7 qui établit un équilibre à ce pH auquel la concentration d'ABA sous sa forme protonée (ABA-H) est égale à la forme anionique non protonée (ABA -). Ainsi, à pH 4,7, 50 % de l'ABA est sous forme d'ABA-H et l'autre 50 % sous forme d'ABA -. Mais le cytosol a un pH plus alcalin (7,2 à 7,4) et l'ABA est principalement sous sa forme anionique ABA- (jusqu'à 1 000 fois plus concentrée que l'ABA-H) et est incapable de diffuser à travers la membrane plasmique. Lorsque le pH diminue (plus acide), la forme protonée, ABA-H, est la forme prédominante, capable de diffuser à travers la membrane sans recourir à un transporteur. Le pH de 5 à 6,1 de l'apoplaste implique qu'entre 40 et 50 % de l'ABA se trouve sous la forme diffusible ABA-H, tandis que de l'autre côté de la membrane plasmique, le pH plus alcalin rend la forme ABA- est la majorité, alors que l'ABA-H ne représente que 0,2 % de l'ABA. Ainsi, dans des conditions normales, la plupart des molécules ABA-H de l'apoplaste qui traversent la membrane plasmique dans le cytosol seront déplacées vers leur forme ionique ABA-. Le cytosol parvient donc à rester avec un déficit constant de la forme ABA-H.
Cependant, lors de situations stressantes, la forte alcalinisation de l'apoplaste ralentirait le processus de simple diffusion de l'ABA vers le cytosol. Ce fait, ajouté à la difficulté de transporter l'ABA du cytosol à l'apoplaste pour son transport vers les cellules cibles, rend nécessaire l'existence de transporteurs. L'identification des transporteurs ABA dans les membranes des cellules cibles, telles que les membranes des cellules occlusives, a résolu le problème de la manière dont l'ABA parvient à pénétrer dans les cellules lorsque le transport passif décroît dans des conditions de stress. L'un des importateurs d'ABA est l'ABCG40. L'expression de l'ABCG40 n'est pas spécifique à un tissu et son produit est situé dans les membranes cellulaires. D'autres importateurs tels que le code ABCG22 pour les transporteurs qui s'expriment principalement dans des cellules occlusives. La sortie de l'ABA dans l'espace intercellulaire est également assurée par des transporteurs tels que ABCG25, qui s'exprime principalement dans les tissus vasculaires, principal site de synthèse de l'ABA.
Rôle de l'ABA dans l'évolution des plantes
Malgré son rôle extrêmement important et connu chez les plantes supérieures, l'ABA n'est pas un composé spécifique de ce groupe mais est présent dans un large éventail d'organismes, des procaryotes aux animaux en passant par les champignons, les protozoaires et les algues. Ce fait suggère immédiatement une origine ancestrale de l'ABA et une fonction plus ou moins conservée en tant que modulateur de la réponse cellulaire aux signaux environnementaux.
Les divers composants qui constituent le coeur de la voie de signalisation ABA chez Arabidopsis (récepteurs PYR / PYL / RCAR, phosphatases du groupe A 2 (PP2C) et kinases SnRK2) ont été conservés dans les plantes terrestres et auraient joué un rôle clé dans la colonisation de l'environnement terrestre par les plantes, en développant un réseau complexe complexe de signalisation pour survivre aux limitations d'eau dans les habitats non aquatiques.
En rapport avec "acide abscissique"
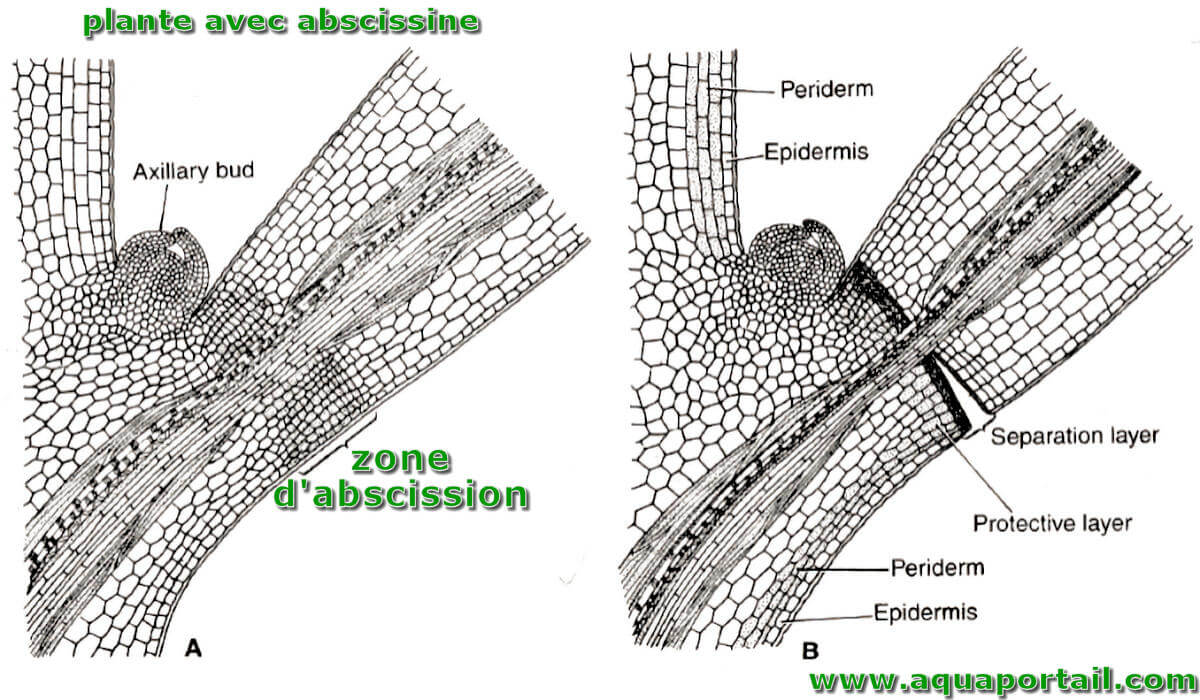
Une abscissine est l'acide d'une phytohormone (acide abscissique) responsable de l'abscission chez les végétaux à feuilles et fleurs.

L'abscission est le processus par lequel les composants végétaux, tels que les fleurs, les fruits et les feuilles, se séparent naturellement de la plante mère.

L'abscission foliaire est un processus préparatoire à la défoliation pour la chute des feuilles.

La zone d'abscission est la zone de séparation cellulaire chez les arbres à feuilles caduques située à la base du pétiole.