
Stomate
Définition
Les stomates sont des structures constituées par un ensemble de cellules situées notamment dans l'épiderme inférieur des feuilles, avec pour fonction d'établir la communication de l'environnement interne avec l'atmosphère, constituant un canal d'échange des gaz et de transpiration du végétal.
Un stomate d'une feuille de tomate :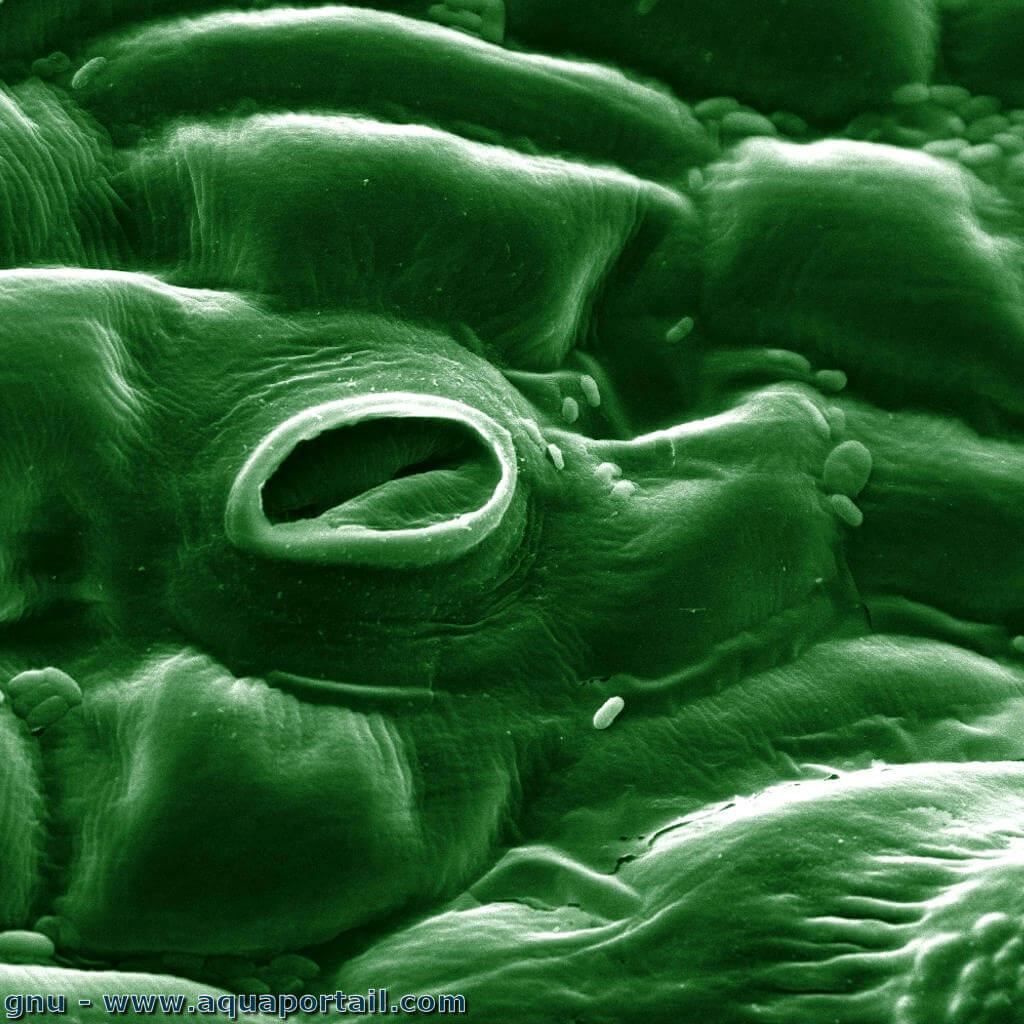
Le stomate, sous forme d'une bouche, dans une feuille de tomate est montré par une image de microscope à balayage électronique colorisé.
En botanique, un stomate est constitué de pores minuscules, situés à la surface des feuilles et des tiges herbacées à travers lesquelles s'opèrent des échanges gazeux avec l'atmosphère. Cette structure épidermique impliquée dans les échanges d'eau et de gaz s'ouvre en fonction des heures de la journée.
Explications
Chez les végétaux supérieurs, les stomates, ou stomas, sont des organes différenciés à la face inférieure (abaxiale) des feuilles et qui sont impliqués dans les échanges gazeux : c'est une feuille hypostomatique.
En milieu aquatique, les plantes aquatiques sont leurs stomates sur la face supérieure (adaxiale) : c'est une feuille épistomatique. Lorsque les stomates se situent sur les deux côtés des feuilles, comme chez les succulentes ou le maïs, c'est une feuille amphistomatique.
En botanique, les deux cellules occlusives qui font partie de l'épiderme de la plante sont dénommées stomates et elles délimitent entre elles un pore appelé ostiole, l'épiderme est imperméable à l'eau et aux gaz grâce à la couche de cuticules que le couvertures, qui permettent de communiquer l'environnement gazeux de l'intérieur de la plante avec celui de l'extérieur.
Ces cellules ainsi que les cellules accompagnant l'épiderme autour d'eux, le cas échéant, forment l'organe appelé dispositif stomatique, qui débouche normalement dans une cavité sous l'épiderme appelée chambre substomatique dont la fonction est d'abriter les gaz.
Les cellules occlusives ouvrent et ferment l'ostiole d'une manière qui est couplée aux conditions physiologiques de la plante, ce qui différencie la stomie d'un pore commun, comme on peut le trouver dans les mousses plus primitives des embryophytes (plantes terrestres).
Une cellule cyclocytique se dit d'un stomate qui est entouré de plusieurs cellules annexes, formant comme un anneau autour de lui.
Un organe syndétochéilique se dit des stomates dont les cellules annexes dérivent de la même cellule mère que les cellules de garde. On les rencontre chez les Chlamydospermes et certaines familles d'Angiospermes.
Fonctions
Les stomates sont localisés dans toutes les parties herbacées ("plantes vertes") des plantes vasculaires et de certaines mousses (c'est-à-dire dans la plupart des plantes terrestres) qui sont les parties dont le tissu protecteur est l'épiderme, sans croissance secondaire.
Avec d'autres organes de fonction similaire (tels que les lenticelles, les zones avec l'aspect verruqueux dans l'écorce des parties ligneuses dont les cellules sont séparées par de grands espaces intercellulaires, donc fonctionnellement des pores), sont la façon dont la plupart des d'oxygène et de dioxyde de carbone, deux gaz utilisés par les cellules à l'intérieur de la plante pendant la photosynthèse et la respiration cellulaire. Ils sont également la principale voie par laquelle la plante perd l'eau absorbée par les racines sous forme de vapeur l'eau (processus appelé transpiration des plantes).
Comme l'échange de gaz est positif mais que la perte d'eau dans cette proportion n'est pas souhaitable, l'évolution a développé deux stratégies qui limitent la perte d'eau à travers les pores. L'un est son emplacement de préférence sur la face inférieure des feuilles, à l'abri du soleil direct, et l'autre est l'évolution des "stomates" avec la possibilité d'ouvrir ou de fermer les pores en fonction des conditions physiologiques de la plante.
Anatomie
Anatomiquement, c'est-à-dire au niveau cellulaire au microscope, on observe que dans tous les stomates il y a deux cellules qui changent de taille en régulant la taille du pore qui, comme sur la figure, le délimite en se réunissant aux deux extrémités. Ces cellules ont été appelées cellules de garde ou cellules occlusives et le pore "ostiole".
Par un signal physiologique, les cellules occlusives peuvent perdre leur rigidité de sorte que lorsque les parois du pore deviennent flasques, elles se referment et se referment, et par le signal physiologique opposé elles augmentent leur turgescence (elles grossissent jusqu'à atteindre la forme de leurs parois cellulaires), qui élargit la cellule dans le sens de la longueur, de sorte qu'elle s'incurve autour du pore, l'ouvrant.
La façon dont les cellules occlusives sont courbées en augmentant leur turgescence est déterminée par la morphologie de leurs parois cellulaires, qui peut varier entre les taxons.
Physiologiquement, les cellules de garde réguleraient leur turgescence en contrôlant la quantité de sels de potassium K+ dans la cellule, en fonction de la quantité de sels à l'extérieur, de sorte que l'eau traverse la membrane cellulaire à l'intérieur ou à l'extérieur de la cellule par osmose, c'est-à-dire à la recherche de la plus forte concentration de solutés.
La situation est régulée par des structures présentes dans la membrane cellulaire qui transfèrent activement les sels et autres ions avec la dépense énergétique, ce que l'on appelle le transport actif et qui est dans cette régulation ionique dans laquelle les cellules d'accompagnement aideraient, le cas échéant.
Les cellules d'accompagnement, ou subsidiaires ou annexées, peuvent ou non être présentes, et sont des cellules épidermiques contiguës aux cellules de garde, avec une morphologie qui les distingue du reste de l'épiderme.
Toutes les plantes terrestres sont recouvertes d'une couche cireuse sécrétée par l'épiderme qui les rend presque imperméables (empêche la perte d'eau), appelée cuticule. La cuticule peut être plus épaisse, comme pour les plantes dans les environnements xérophytiques, ou plus mince, comme pour les plantes dans les environnements humides.
L'épaisseur peut également dépendre des conditions environnementales dans lesquelles la plante a grandi et peut être différente dans différents organes verts de la plante mais il n'est interrompu que dans l'ostiole des stomates.
Cette couche de cire, non présente chez les algues aquatiques dont les stomates sont issus, était nécessaire à leur transition vers la vie terrestre, car elle empêche le séchage extrême qu'elles subiraient si elles ne l'avaient pas. Elle empêche également la plante de capturer et libérer les gaz utilisés ou mis au rebut dans la photosynthèse et la respiration, de sorte que parallèlement à la couche de cire, différents types de pores ont évolué dans les plantes, ainsi que l'environnement interne qui abrite une concentration de gaz qui permet les activités chimiques de la plante.
Les premiers pores apparus dans les plantes terrestres étaient de morphologie simple et on observe encore ces "pores" de morphologie simple dans certaines mousses, qui peuvent avoir une fine cuticule et ne sont pas si propres à résister à la dessiccation.
Lorsque les plantes vasculaires ou trachéophytes (un groupe qui comprend des fougères et des plantes avec des graines) naissent, ou peut-être un peu plus tôt, lorsque les mousses se reproduisent avec elles, la couche de cire devient plus épaisse et les "stomates" de la morphologie plus complexe.
L'anatomie de plusieurs stomates :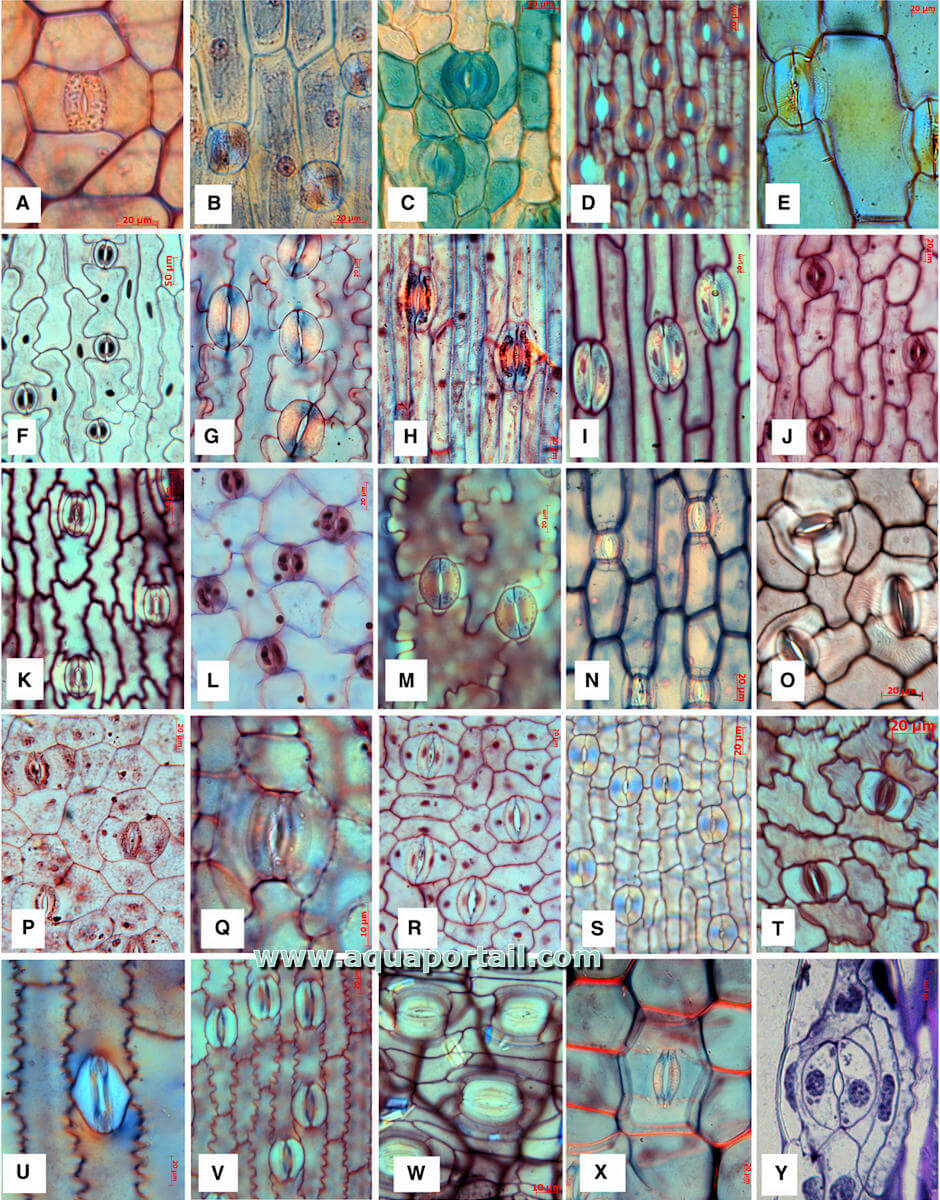
Stomates matures chez les monocotylédones. (A) Astelia menziesii (Asteliaceae – Asparagales). (B) Ixiolirion tataricum (Ixioliriaceae – Asparagales). (C) Diplarrhena moraea (Iridaceae – Asparagales). (D) Aphyllanthes monspeliensis (Asparagaceae s.l. – Asparagales). (E) Dactylorhiza praetermissa (Orchidaceae – Asparagales). (F) Erythronium americanum (Liliaceae − Liliales). (G) Cardiocrinum giganteum (Liliacées – Liliales). (H) Campynema lineare (Campynemataceae – Liliales). (I) Fritillaria pyrenaica (Liliacées – Liliales). (J) Smilax trifolia (Smilacaceae – Liliales). (K) Heloniopsis japonica (Melanthiaceae – Liliales). (L) Luzuriaga radicans (Alstroemeriaceae – Liliales). (M) Drymophila cyanocarpa (Alstroemeriaceae – Liliales). (N) Aletris pauciflora (Nartheciaceae – Dioscoreales). (O) Dioscorea bulbifera (Dioscoreaceae – Dioscoreales). (P) Cyclanthus bipartitus (Cyclanthacées – Pandanales). (Q) Vellozia elegans (Velloziaceae – Pandanales). (R) Damasonium stellatum (Alismataceae – Alismatales). (S) Tofieldia pusilla (Tofieldiaceae – Alismatales). (T) Aglaonema augustifolium (Araceae – Alismatales). (U) Zea mays (Poacées – Poales). (V) carex viridula (Cyperaceae − Poales). (W) Musa paradisiaca (Musaceae – Zingiberales). (X) Zebrina pendula (Commelinaceae – Commelinales). (Y) Borassus flabellifer (Arecaceae – Arecales). Toutes les images orientées avec l'axe principal de la feuille allant de haut en bas.
Les stomates sont l'un des acteurs de la photosynthèse et aussi de la respiration des plantes puisqu'ils passent par l'échange gazeux mécanique, c'est-à-dire que l'oxygène sort à cet endroit et que le dioxyde de carbone entre (photosynthèse) et vice versa (pendant la respiration) selon les processus chimiques qui interviennent dans les plantes.
La suberine est synthétisée uniquement dans les organes qui développent le liège (phloème) sous forme de racines, de tiges et de certains fruits.
Physiologie
L'ouverture ou la fermeture des stomates fait partie de la physiologie de la plante qui communique la plante avec les conditions environnementales et l'adapte à celle-ci.
L'ouverture ou la fermeture des stomates est très finement régulée dans la plante par des facteurs environnementaux tels que la lumière, la concentration de dioxyde de carbone ou la disponibilité de l'eau. Selon les recherches, il est connu que certains cations tels que le potassium et le calcium et les anions tels que le chlorure interviennent activement dans l'ouverture et la fermeture des stomates.
En cas de sécheresse donnant un stress de nature hydrique (stress hydrique), les stomates se ferment en empêchant les pertes d'eau dans l'installation, ce qui empêche également l'échange de gaz et, par conséquent, l'entrée de dioxyde de carbone atmosphérique (gaz carbonique CO2) nécessaire pour la nutrition des plantes à travers le processus de photosynthèse.
C'est pourquoi dans les régions xérophiles, les stomates sont souvent petits ou presque inexistants, et contiennent également des quantités appréciables de cire, de poils et de trichomes, ce qui rend difficile l'évacuation de la vapeur d'eau.
La taille de ceux-ci varie avec le type de stomie qui est observée, mais se situe généralement entre 0,6 et 3,5 micromètres.
Développement
Les stomates commencent leur développement quand une cellule mère, méristématique, subit une division asymétrique, donnant lieu à une méristémoïde et à une plus grande cellule soeur.
À son tour, le méristémoïde (voir méristème) peut continuer à effectuer des divisions asymétriques ou il peut être différencié jusqu'à ce qu'il devienne une cellule de garde mère. Enfin, la cellule de garde mère subit une division symétrique et se différencie en une paire de cellules de garde matures autour d'un pore stomatique.
Les cellules stomatiques ont une paroi interne élastique et plus épais que l'externe, une caractéristique importante du mécanisme d'ouverture et de fermeture des stomates.
Types de stomates
Les taxonomistes ont observé au microscope les stomates de nombreuses espèces végétales et ont observé que les espèces sont regroupées selon leur "type de stomie" dans les groupes de parenté qui ont traditionnellement été placés dans la catégorie de la famille.
Le "type de stomate" est un caractère de valeur taxonomique quand on veut découvrir la parenté d'un taxon au niveau de cette catégorie taxonomique, et très probablement ne sert pas, par exemple, à différencier une espèce d'une autre proche, puisqu'ils partagent probablement le même type de stomie.
Les types de stomie suivants sont pertinents pour le taxonomiste des plantes et seulement avec les figures qui l'accompagnent qui indiquent clairement ce qu'ils doivent observer sous un microscope :
- Anomocitique : il ne possède pas de cellules attachées, il est le plus fréquent chez les dicotylédones et aussi les plus anciennes. Par exemple, les renonculacées Ranunculaceae.
- Paracitique : il a 2 cellules attachées, disposées parallèlement aux occlusales. Par exemple, les Rubiacées Rubiaceae.
- Anisocitique : il a 3 cellules attachées, 1 plus petit. Par exemple, des crucifères Brassicacées Brassicaceae, les Solanacées Solanaceae.
- Tétracytique : il a 4 cellules subsidiaires. Commun dans plusieurs familles de monocotylédones telles que les Aracées Araceae, les Commélinacées Commelinaceae, les musacées Musaceae.
- Diacétique ou caryophyllienne : il possède 2 cellules attachées perpendiculairement à l'occlusif. Peu de familles, les Caryophyllacées, les Acanthacées.
- Cyclocytique : il a de nombreuses cellules subsidiaires, disposées en un ou deux cercles autour des cellules occlusives.
- Hélicocytique : il a plusieurs cellules subsidiaires arrangées dans une spirale autour des arrêts.
Un organe haplochéilique détermine des stomates qui sont issus de la division d'une cellule mère unique. On les rencontre chez les Cycadales, les Ginkgoales, les Ephédrales, les Renonculacées et les Monocotylédones. On distingue :
- les stomates haplochéiliques monocycliques lorsqu'ils ne sont entourés que par deux cellules annexes;
- les stomates haplochéiliques amphicycliques lorsque les cellules annexes sont elles-mêmes entourées par des cellules issues de leur propre division.
Haplochéilique
Un organe haplochéilique détermine des stomates qui sont issus de la division d'une cellule mère unique. On les rencontre chez les Cycadales, les Ginkgoales, les Ephédrales, les Renonculacées et les Monocotylédones. Les stomates ont le plus souvent une structure caractéristique, dite haplochéilique.
On distingue :
- les stomates haplochéiliques monocycliques lorsqu'ils ne sont entourés que par deux cellules annexes;
- les stomates haplochéiliques amphicycliques lorsque les cellules annexes sont elles-mêmes entourées par des cellules issues de leur propre division.
La disposition des stomates chez les Bennettitales (cycadéoïdes) est décrite comme syndétochéilique, c'est-à-dire que les cellules de garde et les cellules subsidiaires se sont développées à partir d'une seule cellule initiale, ce qui donne une disposition très différente de celle des Cycadales, qui ont une disposition haplochéilique similaire à celle des conifères, où deux les cellules de garde sont entourées d'un anneau de cellules subsidiaires.
L'arrangement différent des stomates est l'un des principaux moyens de distinguer les fossiles de feuillage de cycas et de Bennettitales lorsque la cuticule est préservée.
Les Bennettitales sont un ordre éteint de plantes à graines qui est apparu pour la première fois au Permien et s'est éteint dans la plupart des régions vers la fin du crétacé.
Synonymes, antonymes
Voir tous les synonymes pour "stomate".1 synonyme (sens proche) de "stomate" :
- stoma
0 antonyme (sens contraire).
Les mots ou les expressions apparentés à STOMATE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
Le mot STOMATE est dans la page 5 des mots en S du lexique du dictionnaire.
Mots en S à proximité
stirpe stockage dans les dépressions du sol stolon stolonifère stomachique stomatestomatelle stomatiquestomatocyte stomochordestomodéum
En rapport avec "stomate"

En botanique, un organe anisocytique se dit d'un stomate qui possède trois cellules annexes de taille inégale.
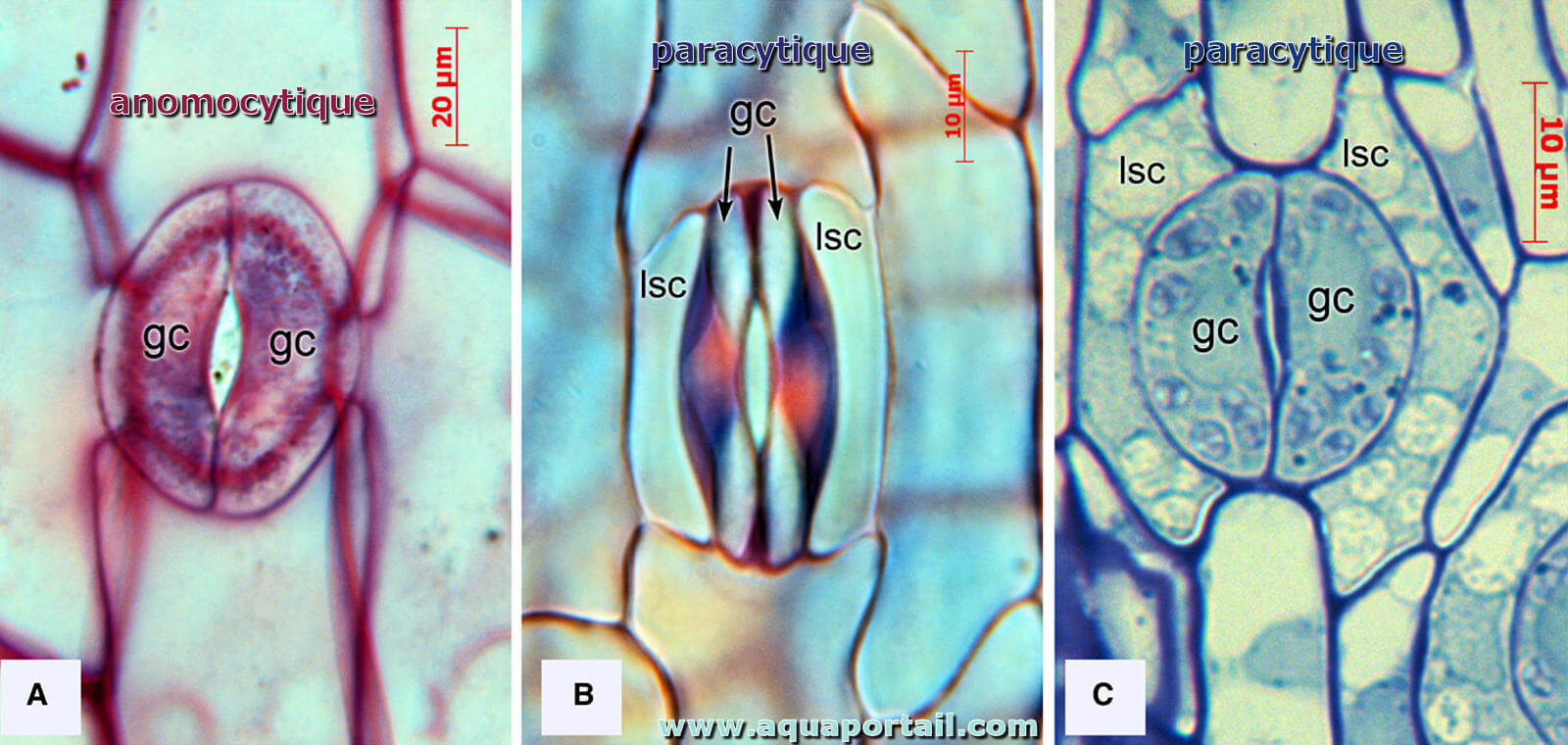
Un organe est anomocytique lorsqu'il qui s'écarte de la norme, anomalistique. En botanique, un stomate anomocytique a une organisation particulière...

La chambre sous-stomatique désigne et identifie une cavité du mésophylle située juste sous un stomate.
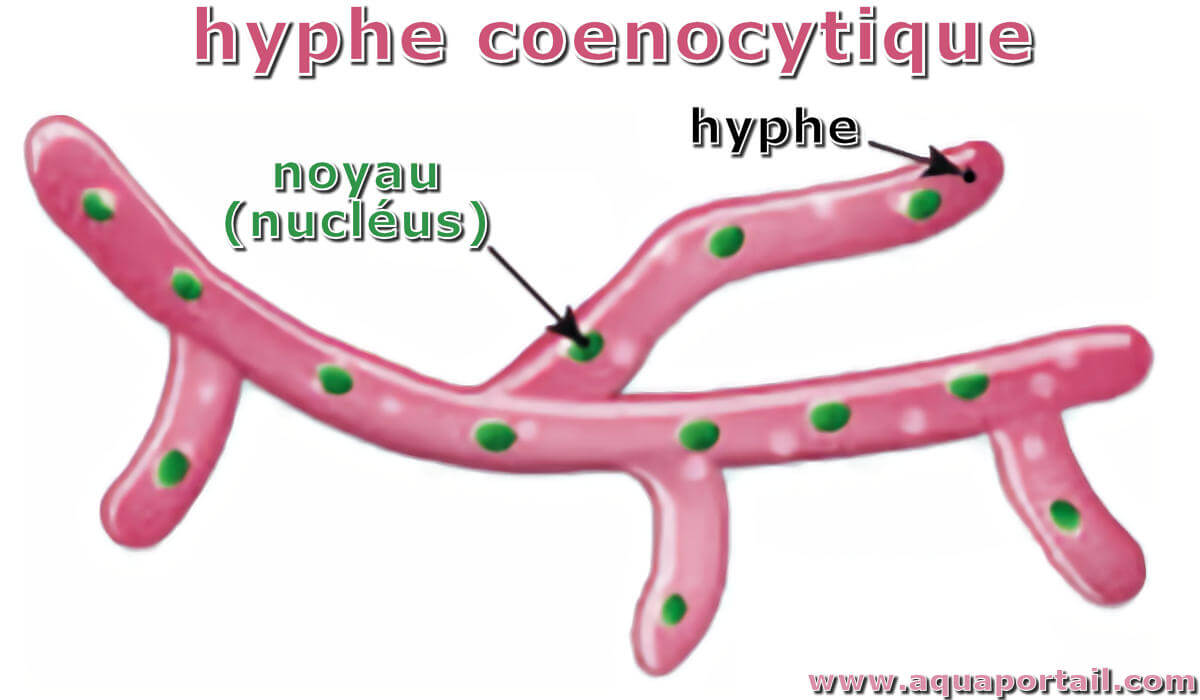
Est coenocytique, ou cénocytique, ce qui présente les caractéristiques d'un coenocyte, souvent pour désigner un hyphe; qualifie le thalle unicellulaire...