
Gluconéogenèse
Définition
La gluconéogenèse (glyconéogenèse, néoglucogenèse, néoglycogenèse) distingue la synthèse de glucose à partir de composés acides non glucidiques comme l'acide gluconique, l'acide pyruvique, l'acide lactique, les intermédiaires du cycle de Krebs, les acides aminés glycoformateurs et le glycérol, seul lipide glycoformateur.
La gluconéogenèse biologique (néoglucogenèse) est stimulée par le glucagon et inhibée par l'insuline. Elle est une biosynthèse pour protéines. Voir aussi la biosynthèse des protéines en génie biologique. Ne pas confondre avec la glycogénogenèse.
Une gluconéogenèse :
Cette gluconéogenèse montre la structure et la genèse du glucose à partir de L-lactate. L'évolution de la gluconéogenèse est répartie chez les eucaryotes sur trois compartiments d'une cellule. La grande majorité se déroule dans le cytosol. Une étape de la réaction existe dans la mitochondrie, une autre dans le réticulum endoplasmique lisse (SER, réticulum endoplasmique lisse anglais), car l'enzyme nécessaire à cet effet (pyruvate carboxylase ou glucose-6-phosphatase) n'est présente qu'ici.
Explications
La gluconéogenèse est une voie métabolique anabolique qui permet la biosynthèse du glucose à partir de précurseurs non glucidiques. Il inclut l'utilisation de plusieurs acides aminés, lactate, pyruvate, glycérol et l'un des intermédiaires du cycle de l'acide tricarboxylique (ou cycle de Krebs) en tant que sources de carbone pour la voie métabolique. Tous les acides aminés, à l'exception de la leucine et de la lysine, peuvent fournir du carbone pour la synthèse du glucose. Les acides gras à chaîne paire ne fournissent pas de carbone pour la synthèse du glucose, car le résultat de leur β-oxydation (acétyl-CoA) n'est pas un substrat gluconéogénique; tandis que les acides gras à chaînes impaires fourniront un squelette carboné qui sera dérivé de l'acétyl-CoA et du succinyl-CoA (qui est un substrat gluconéogénique car c'est un intermédiaire du cycle de Krebs).
Certains tissus, tels que le cerveau, les érythrocytes, les reins, la cornée de l'oeil et des muscles, lorsque l'individu exerce une activité intense, nécessitent un apport continu de glucose, l'obtenir à partir de glycogène hépatique, qui ne peut que satisfaire ces besoins pendant 10 à 18 heures maximum, ce qui prend pour épuiser le glycogène stocké dans le foie. Par la suite, la formation de glucose commence à partir de substrats autres que le glycogène.
La gluconéogenèse existe presque exclusivement dans le foie (10 % dans les reins). C'est un processus clé car il permet aux organismes supérieurs d'obtenir du glucose dans des états métaboliques tels que le jeûne.
Réactions de la gluconéogenèse
Les enzymes qui participent à la voie glycolytique participent également à la gluconéogenèse; Les deux voies se différencient par trois réactions irréversibles utilisant des enzymes spécifiques à ce processus et les deux détours métaboliques de cette voie.
Ces réactions sont :
- Du glucose-6-phosphate au glucose.
- Du fructose-1,6-bisphosphate au fructose-6-phosphate.
- Du pyruvate au phosphoénolpyruvate.
Conversion du pyruvate en phosphoénolpyruvate
L'oxaloacétate est un intermédiaire dans la production de phosphoénolpyruvate dans la gluconéogenèse. La conversion du pyruvate en phosphoénolpyruvate dans la gluconéogenèse est réalisée en deux étapes. Le premier de ceux-ci est la réaction du pyruvate et du dioxyde de carbone pour donner de l'oxaloacétate. Cette étape nécessite de l'énergie disponible par hydrolyse de l'ATP.
L'enzyme qui catalyse cette réaction est la pyruvate carboxylase, une enzyme allostérique trouvée dans les mitochondries. L'acétyl-CoA est un effecteur allostérique qui active la pyruvate carboxylase. Lorsqu'il y a plus d'acétyl-CoA que nécessaire pour maintenir le cycle de l'acide citrique, le pyruvate est dirigé vers la gluconéogenèse. L'ion magnésium et la biotine sont nécessaires pour une catalyse efficace.
La biotine, liée de manière covalente à l'enzyme, réagit avec le CO2, qui se lie par covalence. Une fois le CO2 incorporé au pyruvate, il se forme un oxaloacétate.
La conversion de l'oxaloacétate en phosphoénolpyruvate est catalysée par l'enzyme phosphoénolpyruvate carboxykinase, présente dans la mitochondrie et dans le cytosol. Cette réaction comprend également l'hydrolyse d'un nucléoside triphosphate, dans ce cas le GTP au lieu de l'ATP.
Conversion du fructose-1,6-bisphosphate en fructose-6-phosphate
La réaction phosphofructokinase 1 de la glycolyse est essentiellement irréversible, mais uniquement parce qu'elle est provoquée par le transfert de phosphate de l'ATP. La réaction qui survient dans la gluconéogenèse pour éviter cette étape consiste en une simple réaction hydrolytique, catalysée par la fructose-1,6-bisphosphatase.
L'enzyme à plusieurs sous-unités nécessite la présence de Mg2+ pour son activité et constitue l'un des principaux sites de contrôle régulant la voie globale de la gluconéogenèse. Le fructose-6-phosphate formé dans cette réaction est ensuite soumis à une isomérisation en glucose-6-phosphate par l'action de la phospho-gluco-isomérase.
Conversion du glucose-6-phosphate en glucose
Le glucose-6-phosphate ne peut pas être converti en glucose par l'action inverse de l'hexokinase ou de la glucokinase; le transfert de phosphate de l'ATP rend la réaction pratiquement irréversible. Une autre enzyme spécifique de la gluconéogenèse, la glucose-6-phosphatase, qui nécessite également du Mg2+, est celle qui entre à la place. Cette réaction de dérivation intervient également par simple hydrolyse.
La glucose-6-phosphatase se trouve principalement dans le réticulum endoplasmique du foie avec son site actif sur le côté luminal (du RE). L'importance de sa localisation dans le foie réside dans le fait qu'une des fonctions du foie est de synthétiser le glucose pour l'exporter vers les tissus par la circulation sanguine.
Régulation
La régulation de la gluconéogenèse est cruciale pour de nombreuses fonctions physiologiques, mais surtout pour le bon fonctionnement du tissu nerveux. Le flux traversant la voie doit augmenter ou diminuer en fonction du lactate produit par les muscles, du glucose contenu dans l'alimentation ou d'autres précurseurs gluconéogènes.
La gluconéogenèse est largement contrôlée par l'alimentation. Les animaux qui ingèrent des glucides en abondance ont de faibles taux de gluconéogenèse, tandis que les animaux à jeun ou ceux qui mangent peu de glucides ont un débit élevé par cette voie.
Comme la gluconéogenèse synthétise le glucose et que la glycolyse le catabolise, il est évident que la gluconéogenèse et la glycolyse doivent être contrôlées réciproquement. En d'autres termes, les conditions intracellulaires qui activent une voie tendent à inhiber l'autre.
Il peut y avoir :
- Régulation par niveaux d'énergie : La fructose 1,6-bisphosphatase est inhibée par des concentrations élevées d'AMP, associées à un état pauvre en énergie. C'est-à-dire que la forte concentration d'AMP et la réduction de l'ATP inhibent la gluconéogenèse.
- Régulation par le fructose 2,6-bisphosphate : La fructose 1,6-bisphosphatase est inhibée par le fructose 2,6-bisphosphate, un modulateur allostérique dont la concentration est déterminée par la concentration sanguine en circulation de glucagon; La fructose 1,6-bisphosphatase est présente dans le foie et les reins.
- Régulation de la phosphorylation : Ce processus dépend de la concentration en ATP; en diminuant la concentration en ATP, on observe également une diminution de la phosphorylation et inversement. Dans le foie, ce processus augmente avec l'augmentation de la synthèse de la glucokinase, processus favorisé par l'insuline. La membrane des hépatocytes est très perméable au glucose, dans le muscle et l'insuline du tissu adipeux agit sur la membrane pour la rendre perméable.
- Régulation allostérique : Le jeûne prolongé augmente l'acétyl-CoA, ce qui stimule la pyruvate carboxylase et donc la gluconéogenèse, tout en inhibant la pyruvate déshydrogénase; l'élévation d'alanine et de glutamine stimule la gluconéogenèse. Le cortisol augmente la disponibilité du substrat et le fructose 2,6-bisphosphate inhibe la fructose 1,6-bisphosphatase.
Bilan énergétique
Il est souligné que les voies cataboliques génèrent de l'énergie, tandis que les voies anaboliques impliquent un coût énergétique. Dans le cas de la gluconéogenèse, nous pouvons calculer ce coût; La synthèse du glucose est coûteuse pour la cellule au sens énergétique. Si nous partons du pyruvate, six groupes phosphates à haute énergie sont consommés, 4 ATP (en raison des réactions de la pyruvate carboxylase et de la phosphoglycérate kinase) et 2 GTP (conséquence de la décarboxylation de l'oxaloacétate), ainsi que 2 du NADH, qui est le équivalent énergétique d'un autre 5 ATP (étant donné que l'oxydation mitochondriale de 1 NADH génère 2,5 ATP).
Par contre, si la glycolyse pouvait agir dans le sens opposé, la dépense énergétique serait beaucoup plus faible : 2 NADH et 2 ATP.
Importance biomédicale
La gluconéogenèse répond aux besoins en glucose du corps lorsqu'elle n'est pas disponible en quantité suffisante dans le régime alimentaire. Un apport constant de glucose est nécessaire en tant que source d'énergie pour le système nerveux et les érythrocytes. De plus, le glucose est le seul carburant qui fournit de l'énergie au muscle squelettique dans des conditions d'anaérobiose. Le glucose est un précurseur du sucre du lait (lactose) dans la glande mammaire et est activement capturé par le bébé.
D'autre part, les mécanismes gluconéogènes sont utilisés pour purifier du sang les produits du métabolisme d'autres tissus. par exemple, le lactate, produit par les muscles et les érythrocytes, et le glycérol, formé en continu par le tissu adipeux.
Gluconéogenèse et glycolyse
La gluconéogenèse ne correspond que partiellement à la réaction inverse de la glycolyse. Cependant, il existe trois réactions dans lesquelles l'équilibre chimique repose presque exclusivement sur les produits de la réaction. Ces étapes, toutes catalysées par des kinases, sont les suivantes :
- la conversion du glucose en glucose-6-phosphate,
- de fructose-6-phosphate dans du fructose-1,6-bisphosphate et
- la réaction du phosphoénolpyruvate (PEP) en pyruvate.
Pour inverser ces réactions, il faudrait que la cellule puisse établir des rapports de concentration extrêmes. Par conséquent, ces trois étapes de la glycolyse sont de facto irréversibles et sont contournées dans la gluconéogenèse dans l'ordre inverse, comme suit :
- la carboxylation du pyruvate en oxaloacétate sous la consommation d'ATP (pyruvate carboxylase) et la décarboxylation subséquente par phosphorylation de l'oxaloacétate en PEP sous la consommation de GTP (phosphoénolpyruvate carboxykinase);
- la fructose-1,6-bisphosphatase catalyse la réaction du fructose-1,6-bisphosphate en fructose-6-phosphate;
- le glucose-6-phosphate est converti en glucose par la glucose-6-phosphatase (dans la glycolyse, une hexokinase ou une glucokinase (hexokinase IV) catalyse la réaction inverse).
Les autres processus de transformation sont en équilibre, raison pour laquelle ils jouent également un rôle dans la gluconéogenèse.
Une autre différence importante par rapport à la glycolyse est le site de réaction. Bien que cela intervienne exclusivement dans le cytosol, la gluconéogenèse est répartie sur trois compartiments. La conversion du pyruvate en oxaloacétate a lieu dans la lumière de la mitochondrie. L'oxaloacétate, cependant, ne peut pas passer librement à travers la membrane interne de la mitochondrie et doit d'abord être transformé. Il y a deux façons de le faire. L'oxaloacétate mitochondrial est converti en PEP par une PEP carboxykinase mitochondriale. Le PEP quitte ensuite la mitochondrie par un système spécial de navette anionique. Dans le cytoplasme, le PEP est converti en glucose à la suite de la gluconéogenèse.
La gluconéogenèse et la glycolyse partagent plusieurs réactions enzymatiques, mais sont deux voies métaboliques complètement opposées. Par conséquent, une régulation est nécessaire; elle se déroule à deux endroits :
- dans les réactions de pyruvate à PEP et
- dans la conversion du fructose 1,6-bisphosphate en fructose 6-phosphate.
Première réaction : la pyruvate kinase catalyse la transformation du PEP en pyruvate, qui existe lors de la glycolyse. L'activité de cette enzyme est augmentée par le fructose-1,6-bisphosphate et inhibée par l'ATP et l'alanine. Les enzymes de la gluconéogenèse (pyruvate carboxylase et PEP-carboxykinase) sont activées par l'acétyl-CoA et inhibées par l'ADP. L'ATP étant converti en ADP par hydrolyse, on peut parler de régulation réciproque dans ce type de régulation de deux réactions opposées. Un autre exemple fournit la réaction indiquée sous 2. La phosphofructokinase impliquée dans la glycolyse est stimulée par le fructose-2,6-bisphosphate et l'adénosine monophosphate (AMP), mais est entre autres inhibée par le citrate. Réciproquement, la régulation de la fructose-1,6-bisphosphatase impliquée dans la gluconéogenèse a lieu (activée par le citrate et inhibée par le fructose-2,6-bisphosphate et l'AMP).
Synonymes, antonymes
Voir tous les synonymes pour "gluconéogenèse".3 synonymes (sens proche) de "gluconéogenèse" :
- glyconéogenèse
- néoglucogenèse
- néoglycogenèse
1 antonyme (sens contraire) :
Les mots ou les expressions apparentés à GLUCONÉOGENÈSE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
Le mot GLUCONEOGENESE est dans la page 2 des mots en G du lexique du dictionnaire.
Mots en G à proximité
glucane glucide glucocorticoïde glucokinase glucolyse gluconéogenèsegluconique glucopyranose glucosamine glucosane glucose
En rapport avec "gluconéogenèse"
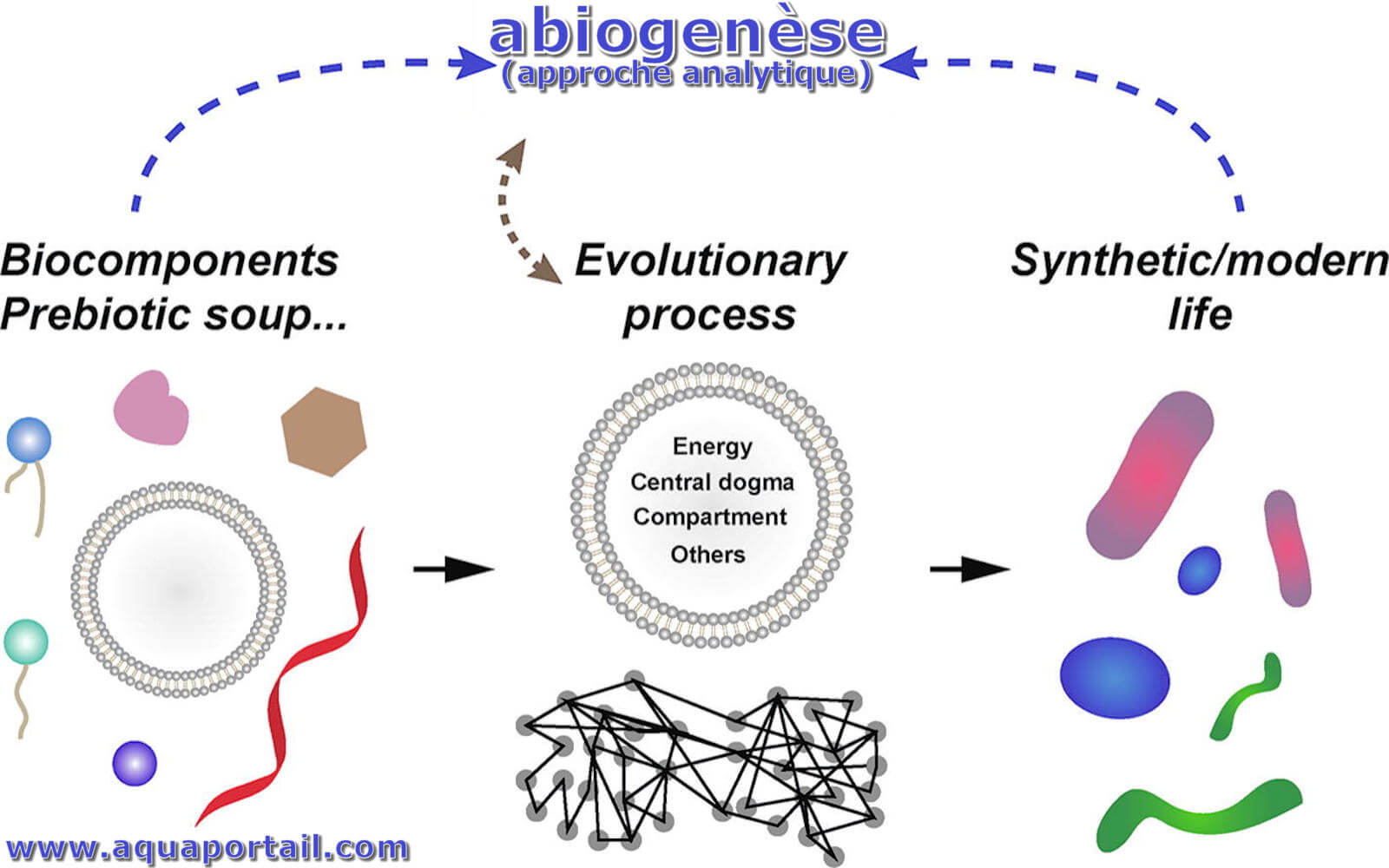
L'abiogenèse est un processus naturel de l'émergence ou de l'origine de la vie à partir de la non-existence de celle-ci, c'est-à-dire à partir de la matière...
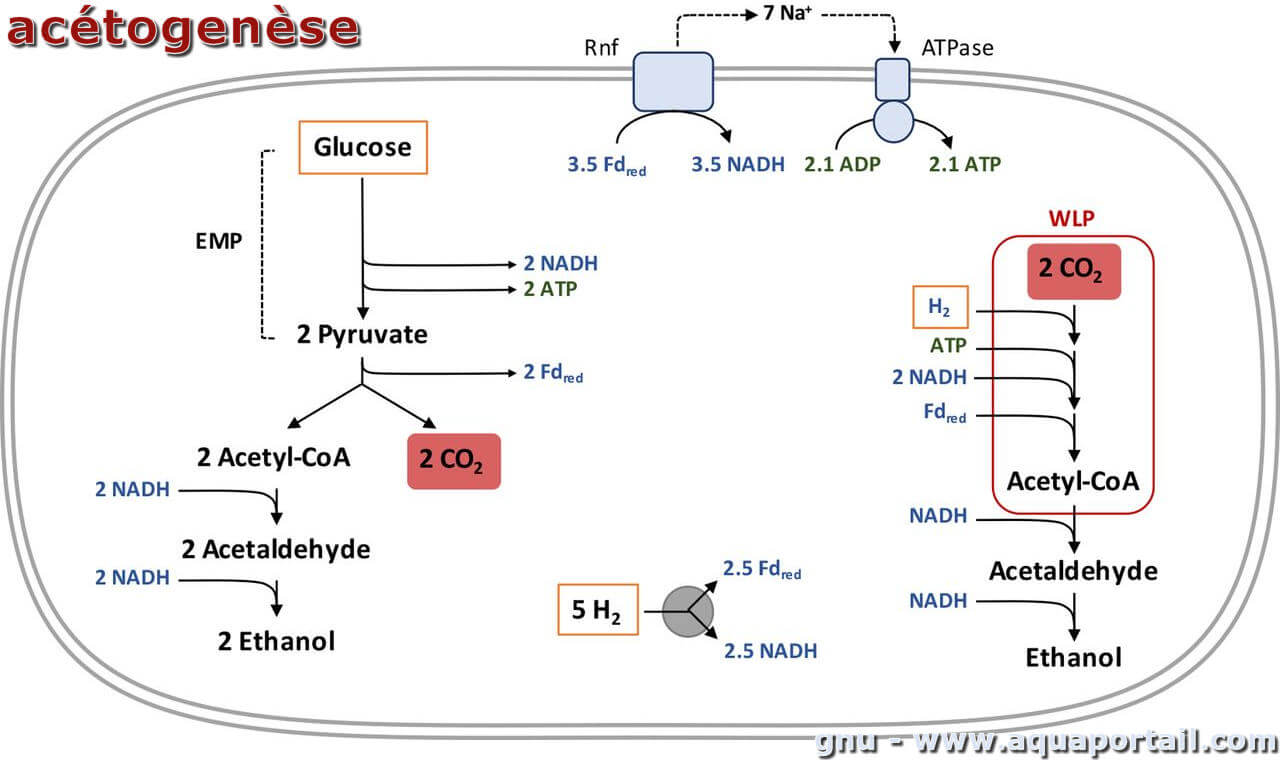
L'acétogenèse est une synthèse bactérienne d'acide acétique (ou acide éthanoïque), par oxydation aérobie de l'éthanol, par fermentation d'hexoses...
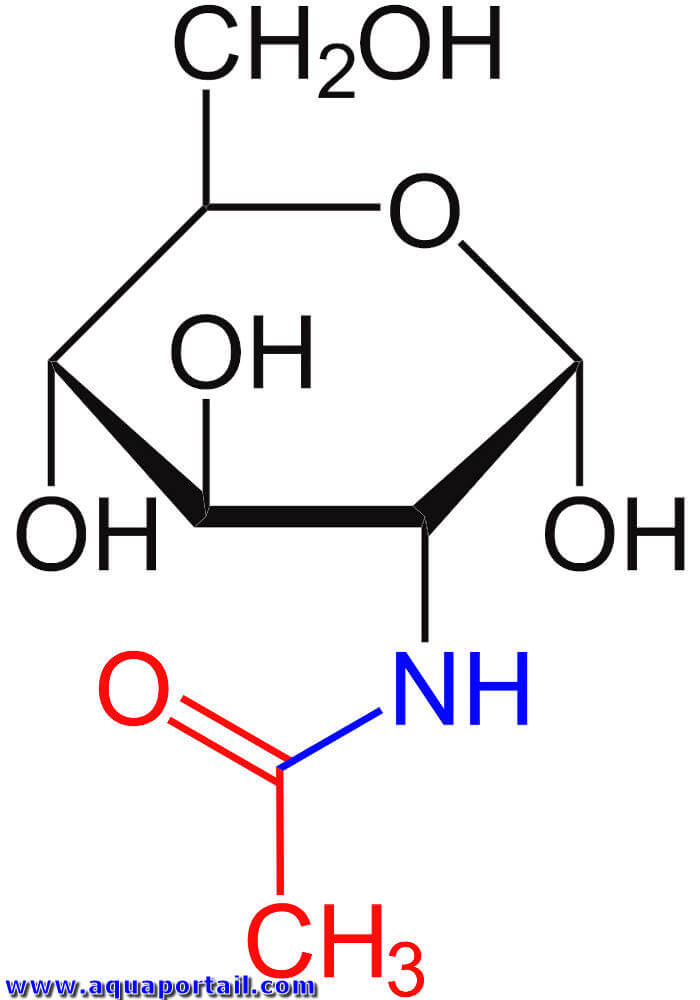
L'acétylglucosamine est un composé de sucre, d'un groupe amine et d'un groupe acétyle, proche de la cellulose.
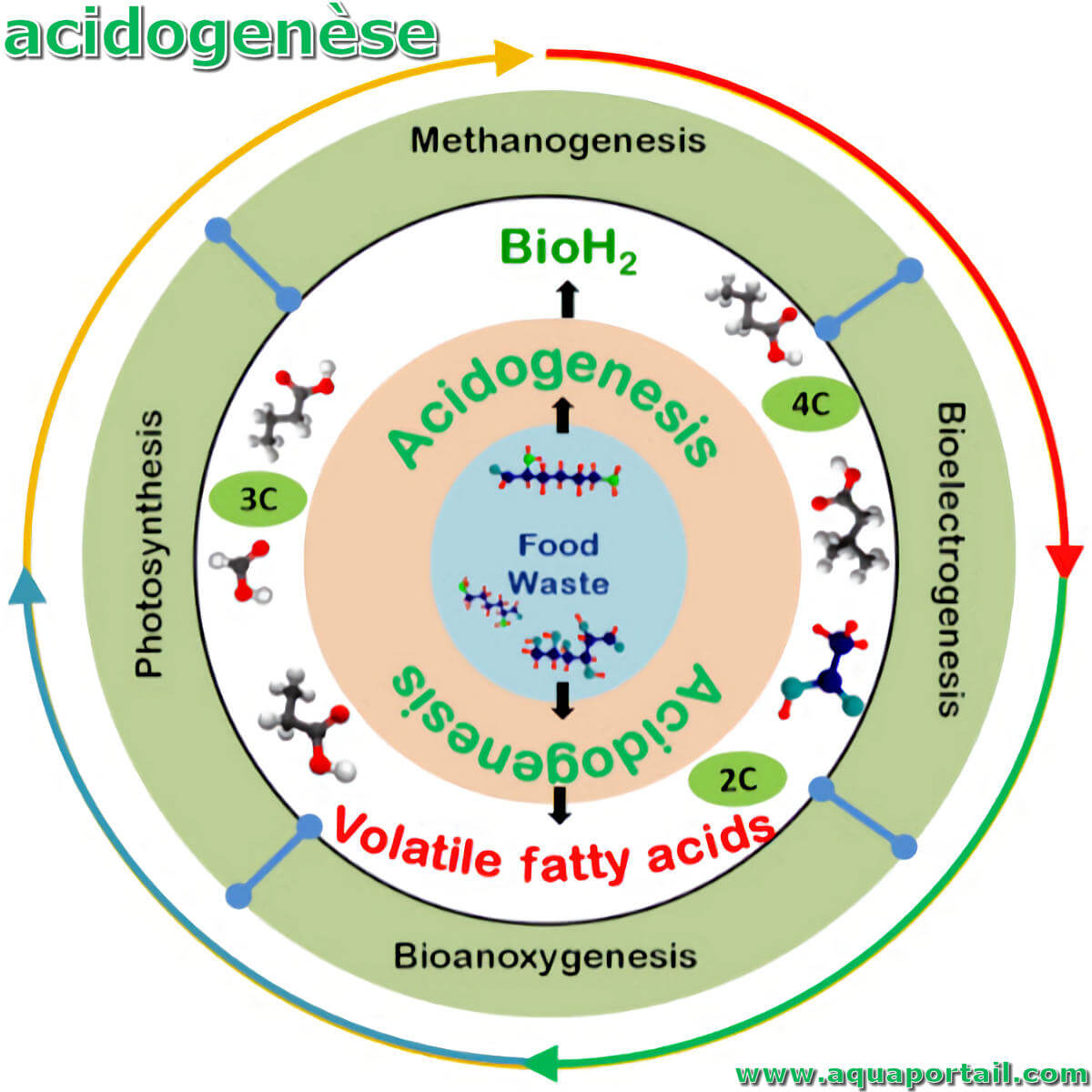
Une acidogenèse représente la deuxième étape dans les quatre étapes de la digestion anaérobie, en précédant l'acétogenèse et la méthanisation.