
Le pollen
Le pollen est une sorte de poudre formée par les grains de pollen libérés par les étamines des plantes à graines. Il constitue l'ensemble de grains microscopiques (comme une farine) produits par les éléments mâles (les étamines) d'un spermatophyte. Le pollen est un gamétophyte mâle de spermaphytes.
Des tubes polliniques relient les grains de pollen :
Le pollen est le nom collectif des grains, plus ou moins microscopiques, produits par des plantes à graines (spermatophytes), contenant chacune un microgamétophyte (gamétophyte mâle). Le sac pollinique est la partie de l'anthère qui contient les grains de pollen, dans les organes mâles de la fleur, les étamines.
Généralités
Les grains de pollen sont dus à une division de réduction (méiose) haploïde, car leurs cellules n'ont qu'un simple ensemble de chromosomes. Ils correspondent aux gamétophytes des mousses et des fougères (voir diplohaplonte, les végétaux diplohaplontiques).
La palynologie est la discipline qui étudie le pollen, par exemple pour l'exine et l'intine, ce qui est pollinique, en particulier sa morphologie et celle du tube pollinique.
Les grains de pollen sont entourés d'une paroi résistante, le sporoderme, constitué de sporopollénine. Ils servent à amener les gamètes (cellules germinales) mâles protégés aux organes récepteurs de la femme et permettent ainsi la pollinisation et, par la suite, la fécondation.
Le pollen est la principale source de nourriture des insectes butineurs et pollinivores (voire nectarivores), notamment les abeilles, dont il est la source essentielle de protéines. Le pollen entre dans la composition de la gelée royale et des miels.
Le pollen peut se composer d'un grain isolé (monade inaperturé, poré, colpé ou colporé) ou de grains multiples (dyade, tétrade, polyade).
Le grain de pollen contient un individu mâle réduit à deux ou trois cellules : le gamétophyte mâle, la phase haploïde dans le cycle d'alternance des générations caractéristiques des plantes. Une fois que la pollinisation a eu lieu, une fois que le grain de pollen arrive à la surface réceptive dans la plante de destination, c'est-à-dire le stigmate, la germination a lieu.
Du grain naît le tube pollinique, qui est une émanation du cytoplasme à travers laquelle migrent les noyaux mâles en direction de l'oosphère (le gamète femelle) et le noyau polaire (chez les Angiospermes, il y a une double fécondation). Voir la microsporogenèse et la microgamétogenèse.
Le grain de pollen a des couches dans sa paroi cellulaire. Le premier est de nature pectocellulosique et ne diffère pas essentiellement de la paroi primaire des cellules somatiques, le second est composé de sporopollénine, un polyterpène qui, grâce à sa structure chimique à forte teneur en carbone, forme un composé résistant à la destruction de surface et permet de préserver les caractéristiques structurelles des grains de pollen.
C'est la base de la palynologie pour atteindre ses déterminations taxonomiques. De plus, chaque type de pollen a une morphologie caractéristique au niveau des espèces, du genre ou de la famille, qui peut avoir des coupes (ouverture dans la coque de grains de pollen), des pores ou les deux.
Le pollen (jaune) d'une fleur de tulipe :
L'anthère d'une fleur de tulipe porte beaucoup de grains de pollen.
En botanique, ce qui est pollinifère signifie qui porte du pollen, ou qui produit du pollen. La plupart des plantes produisant des fleurs sont pollinifères car ces plantes à fleurs produisent des pollinies. Les animaux pollinifères les plus connus sont des insectes comme les abeilles. Tout arum, par exemple l'arum des marais, est pollinifère.
Voir aussi la pollination, et la pollinie pour le cas spécifique des orchidées.
Structure et formation du pollen
Les grains de pollen ont des tailles, formes et structures de surface très diverses et peuvent souvent être attribués en raison de ces caractéristiques de l'espèce ou du moins du genre. La plupart des grains de pollen ont une taille comprise entre 10 et 100 microns, le plus petit étant d'un diamètre de 5 µm chez Myosotis, le plus gros étant chez Cucurbita avec un diamètre de 170 à plus de 200 microns.
Le pollen montre une grande variation dans la taille, la forme et le motif de surface.
Pollen bicellulaire et pollen tricellulaire
Le pollen est la génération de microgamétophytes haploïdes dans les plantes à graines et est considéré comme le partenaire mâle dans la reproduction sexuée. Dans les plantes à fleurs, le pollen comprend deux ou trois cellules lorsqu'il est éliminé de la fleur. Les deux types de pollen possèdent une grande cellule végétative contenant une seule cellule générative (pollen bicellulaire) ou une paire de spermatozoïdes (pollen tricellulaire).
Pollen bicellulaire vs. pollen tricellulaire :
Le pollen bicellulaire (à gauche) est comparé au pollen tricellulaire (à droite).
Environ 30 % des plantes à fleurs ont un pollen tricellulaire contenant des spermatozoïdes entièrement formés à l'anthèse.
La plupart des angiospermes libèrent du pollen bicellulaire. Cependant, dans environ un tiers des angiospermes existants, la deuxième mitose du pollen intervient avant l'anthèse, de sorte que le pollen est tricellulaire lors de sa libération. Le passage du développement bicellulaire au développement tricellulaire s'est produit plusieurs fois indépendamment, mais ses causes sont largement inconnues.
La coexistence des deux types de pollen à la déhiscence des anthères est possible chez quelques espèces. L'examen du nombre de cellules polliniques au cours du développement des anthères a montré que cette coexistence était due à une mitose tardive débutant peu de temps avant l'excrétion du pollen. Les deux types de pollen ont également bien germé au cours du développement.
Étant donné que des proportions variables de pollen bicellulaire et tricellulaire ont été observées à différents moments d'échantillonnage, le rôle de la température a été testé en effectuant des expériences sur le terrain et en chambre de croissance, qui ont montré que des températures plus élevées près de l'anthèse avançaient le temps de la mitose du pollen II.
Les résultats montrent que la sélection pourrait favoriser la production de pollen tricellulaire dans certaines circonstances environnementales qui favorisent la germination rapide du pollen et fournissent la preuve d'un système dans lequel la variation du développement persiste, mais qui peut être modifiée par des facteurs externes tels que la température.
Intine et exine
Les grains de pollen ont une paroi résistante appelée sporoderme. Le sporoderme est constitué de deux couches complexes : l'intine interne et l'exine externe.
L'intine (endospore) est généralement tendre et peu résistante. Souvent, il se compose de deux à trois couches, la plus externe (exospore) ayant une teneur élevée en pectine, permettant une libération facile de l'exine. Les couches internes sont principalement constituées de fibrilles de cellulose. Lorsque le grain de pollen germe, la cellule végétative entourée par l'intestin se transforme en un tube à pollen.
Le composant principal de l'exine est la sporopollénine résistante, qui forme l'exine en gros granules d'environ six nanomètres. Cela se compose de deux couches : l'endexine interne et l'ectexine externe :
- Chez les Gymnospermes, l'endexine a une structure lamellaire. L'ectexine consiste à nouveau en une couche de pied interne et une couche externe compacte, qui enveloppent une couche intermédiaire granuleuse ou alvéolaire.
- Chez les angiospermes, les endexines sont granulaires. Les endexines et la couche de pied dense des ectexines sont combinées en liaisons. Le reste de l'ectexine forme la sexine, qui est généralement très fortement structurée. Si la sexine est constituée de tiges, de trèfles, de cônes, de verrues et de structures similaires, mais sans couche extérieure, elle est appelée grain de pollen ou monade inaperturé. Dans les grains de pollen porés, les colonnes (columelles) à l'extérieur sont reliées pour former une couche, le tectum. Le tectum peut à nouveau être très varié : ouvert, multicouche et sculpté à nouveau. Dans les cavités du tectum, diverses substances sont stockées ou stockées :
- La "colle de pollen" est une substance huileuse de lipides et de caroténoïdes qui fait adhérer les grains de pollen aux pollinisateurs. Cette colle du pollen n'est formée que d'angiospermes, mais peut être absent.
- Des protéines d'incompatibilité qui servent à empêcher l'autofécondation.
La structure de l'exine est déterminée par le grain de pollen. Le matériau, la sporopollénine, est toutefois formé par le tapetum de la paroi de l'anthère et se superpose au grain de pollen.
Ouvertures (apertures)
Les grains de pollen contiennent une ou plusieurs ouvertures germinales. À ces points, l'exine est manquante. À travers l'une des ouvertures, le tube pollinique peut se développer pendant la germination du pollen. Le grain de pollen a un pôle proximal, c'est-à-dire le pôle pointant vers le centre de la tectrade du pollen, et un pôle distal. Le plan équatorial est perpendiculaire aux pôles :
- Le pollen sans ouverture est appelé inaperturé, comme une aperture.
- Les lignes germinales allongées sont appelées sulcus lorsqu'elles se trouvent sur le pôle distal. Le pollen est alors poré. C'est le cas pour la plupart des graines nues.
- Les lignées germinales réparties équatoriales ou sur toute la surface sont appelées colpus, le pollen est colpé.
- Les ouvertures germinales composites sont appelées colporées.
Un pollen tricolpate :
Exemple d'un grain de pollen tricolpate d'une eudicotylédone du genre Arabis (des Brassicacées appelées les arabettes).
Selon le nombre d'ouvertures germinales, une distinction est faite entre mono- (en), tri- (trois), poly- (plus de trois dans le plan équatorial) et pento-aperturé (plus de trois pollens répartis sur toute la surface). Les structures situées dans le plan équatorial sont désignées par le préfixe zono- (zonoporé, zonocolpé...).
Chez les monocotylédones et les dicotylédones primitives (Magnoliidae), le pollen monophore est prédominant. Chez les espèces tricolpates de Rosopsida, le pollen et ses variations sont prédominants.
L'ouverture peut être recouverte d'un opercule, une structure complètement séparée du reste de la sexine.
Charge électrostatique
Le pollen porte souvent des charges d'électricité statique. Étant donné que de nombreuses plantes à fleurs sont capables d'orientation électrique active, la plante peut enregistrer l'arrivée de pollen (adhérant également aux insectes) dans la fleur et ouvrir davantage les fleurs, par exemple.
Le champ électrique qu'elle produit peut changer de plante en quelques secondes pour réagir au pollen ou aux insectes entrants. Mais même avec la pollinisation par le vent, le différentiel de charge du pollen présente un avantage sélectif. Ce phénomène est utilisé pour la pollinisation électrostatique industrielle.
Unités de propagation
Dans la plupart des cas, les grains de pollen sont répartis individuellement, sous forme de monades. En outre, ils peuvent également être déchargés par groupes de deux ou quatre grains dans l'air, on parle alors de dyades ou de tétrades.
Les tétrades se forment lorsque les cellules filles d'une cellule mère de pollen sont collées ensemble. C'est le cas des éricacées Ericaceae. Chez les Cypéracées Cyperaceae, trois des quatre cellules filles sont réduites pour produire des Pseudomonas.
Les grains de pollen peuvent être regroupés pour former des groupes plus importants, et cela par colle à pollen; une autre possibilité est des fils collants de sporopollénine, de la cellulose et/ou des protéines. Si les grains de pollen de plusieurs cellules mères de pollen restent liés, des polyadénoses de 8, 16 ou 32 grains de pollen se forment, par exemple chez les Mimosoideae. De plus grands groupes sont également appelés des massues ou des pollinies, comme par exemple l'acacia et beaucoup d'orchidées.
Si tout le contenu d'un ou de plusieurs sacs polliniques reste cohérent, on l'appelle pollinie. La pollinie est souvent entourée d'une coquille de sporopollénine commune et se rencontre chez certains représentants des Apocynaceae et des orchidées. Le pollinarium est appelé pollinie avec les appendices, qui servent à se propager; dans le cas des orchidées, il s'agit de tiges et de disques collants. La définition du pollinarium est : avec plusieurs sacs à pollen (plusieurs pollinies).
Il existe une corrélation entre la taille des grains de pollen et les pollinisateurs, suite à coévolution. Plus un pollinisateur est spécifiquement adapté à une espèce de plante, telle que les orchidées, plus grands sont les paquets de pollen qui lui sont donnés. Dans le cas de pollinisateurs non spécifiques (par exemple de nombreux Coléoptères), les paquets de pollen individuels sont faibles, car la probabilité que le pollinisateur retourne sur une fleur de la même espèce est faible.
Développement du grain de pollen
Les grains de pollen se forment dans les sacs à pollen des anthères d'une étamine. De chaque cellule mère de pollen (également appelée cellule mère de microspore), quatre grains de pollen haploïde émergent après la méiose. Au cours de la maturation, au moins une autre division cellulaire existe dans chaque grain de pollen, une ou deux petites cellules génératrices et une plus grande cellule végétative environnante apparaissant dans les capillaires.
Le nombre de cellules chez les mammifères nus est plus élevé, jusqu'à un maximum de 50. Les grains de pollen sont nourris par le tapetum, la couche la plus interne de la paroi de l'anthère. Les substances de réserve (protéines, glucides et lipides) sont stockées dans la cellule végétative et l'humidité est extraite du grain de pollen pendant la maturation.
La formation de la paroi du grain de pollen commence au stade unicellulaire avec la formation d'une primexine par la cellule pollinique. Celle-ci est principalement constituée de cellulose et sert de matrice dans laquelle sont ensuite stockées les sporopollénines formées par le tapetum environnant. Après la formation de la primexine, le grain s'entoure de l'intine.
Propagation (dispersion) du pollen
Le pollen est disséminé par le vent (anémogamie), l'eau (hydrogamie) ou les animaux (zoogamie par les insectes ou oiseaux, parfois certains mammifères comme les chauve-souris). La numération pollinique existe depuis environ 300 millions d'années et est indispensable pour plus de la moitié des végétaux à pollinisation. Le pollen peut être transféré d'une fleur à une autre : cas de l'autopollinisation.
La pollination qualifie le moment de l'émission de pollen par le couple étamines-pistils pour la pollinisation, détermine aussi une plante pollinante, un végétal pollinant. La pollination intervient juste avant la pollinisation. Une inflorescence pollinante se dit d'une fleur, d'une plante, d'un peuplement végétal, au moment de la libération du pollen ou pollination.
Science du pollen
Comme le pollen est largement distribué par le vent et reste souvent dans les sédiments lacustres ou les tourbes, il présente un grand intérêt pour la recherche en géologie et sur le climat (climatologie végétale). Sur la base du pollen trouvé, des conclusions peuvent être tirées, par exemple, sur l'histoire de la forêt en Europe centrale et donc également sur le climat d'une période géologique.
La science traitant de ce sujet s'appelle la palynologie. En raison du pollen contenu, l'origine du miel peut être déterminée. Cette tâche s'appelle la mélissopalynologie.
Les analyses de pollen sont également utilisées pour enquêter sur des crimes (palynologie légale). Pour la première fois en 1959, une analyse de pollen élucida un meurtre en Autriche.
Récolte de pollen
En apiculture, un dispositif spécial, appelé piège à pollen, peut être fixé à l'entrée d'une ruche. Il s'agit essentiellement d'une grille à travers laquelle les abeilles qui reviennent doivent se faufiler, perdant ainsi leur "pelote de pollen".
Les masses polliniques (pelotes de pollen) tombent dans un récipient collecteur, qui est généralement vidé deux fois par jour. Ensuite, le pollen ainsi obtenu doit être immédiatement nettoyé (éliminer les corps étrangers) et séché. Comme le pain d'abeille, qui est également composé principalement de pollen, le pollen est comestible.
Le piège à pollen doit être enlevé régulièrement afin de garantir l'approvisionnement en protéines nécessaire à la reproduction du couvain.
Allergies
Le pollen transmis par le vent pose problème à de nombreuses personnes allergiques. Les grains de pollen libèrent un certain nombre de protéines, de lipides et de sucres lors du contact avec une phase aqueuse.
Certaines protéines et certains lipides produisent une réaction immunitaire spécifique qui déclenche une réaction allergique au deuxième contact et à chaque contact ultérieur. Cela peut être accompagné de yeux rouges et larmoyants, d'éternuements et d'écoulement nasal (rhinite allergique). Les concentrations de pollen sont élevées le matin et le soir en ville. La iatropalynologie traite de l'élucidation de ces effets.
Polliniphage
Est polliniphage ce qui se nourrit de pollen, ou dont l'alimentation dépend de la présence de pollen. Le régime phagique, ou régime alimentaire, est, par exemple, adopté par certains pucerons ou leur prédateur, les coccinelles. Le pollen étant considéré comme végétal, l'organisme a une phagie de type phytophage.
Des nombreux insectes volants comme les abeilles, les guêpes, les papillons, sont, au moins partiellement, polliniphages.
En rapport avec "pollen"
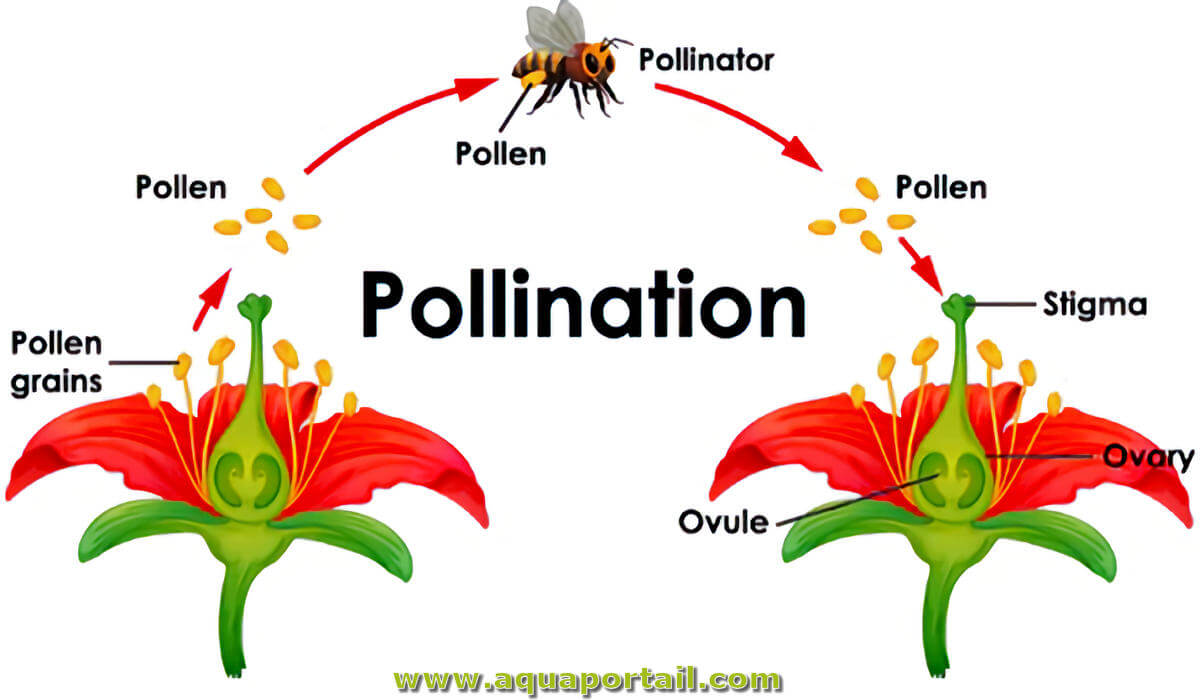
La pollination est l'exact moment du transfert des grains de pollen de l'anthère mâle d'une fleur au stigmate femelle.
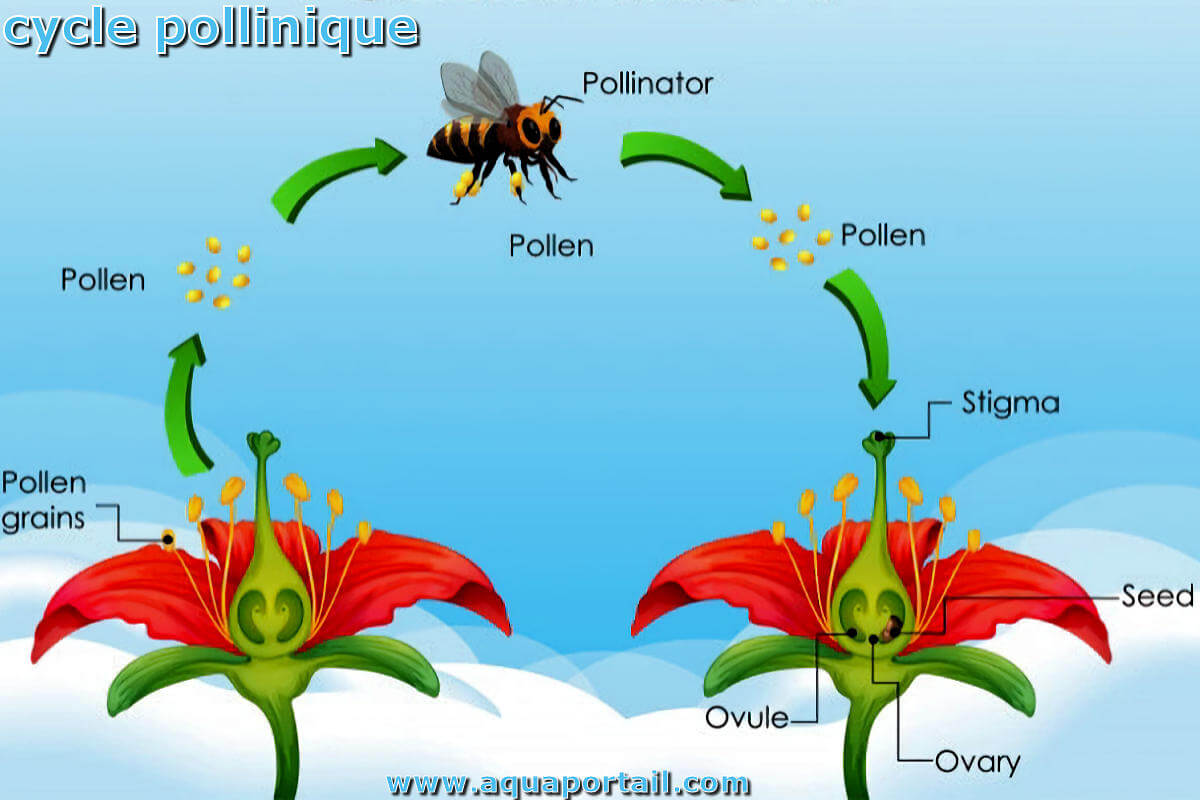
En biologie, est pollinique ce qui est relatif au pollen et à la pollinisation. Tout organe pollinique est en rapport avec le pollen, comme le tube...

Une analyse pollinique est l'ensemble des techniques de détermination et de comptage des spores et pollens d'un sédiment, aux fins de reconstituer...

Une autopollinisation, ou l'autofécondation, précise le mode de pollinisation du pistil d'une fleur par son propre pollen.