
Le système nerveux entérique (SNE)
Le système nerveux entérique (SNE) est la composante du système nerveux autonome formée d'un réseau de neurones et de cellules gliales dans la paroi de l'intestin. Il contrôle la motilité, le flux sanguin, l'absorption de nutriments, la sécrétion et les processus inflammatoires dans le tractus gastro-intestinal.
Le système nerveux entérique :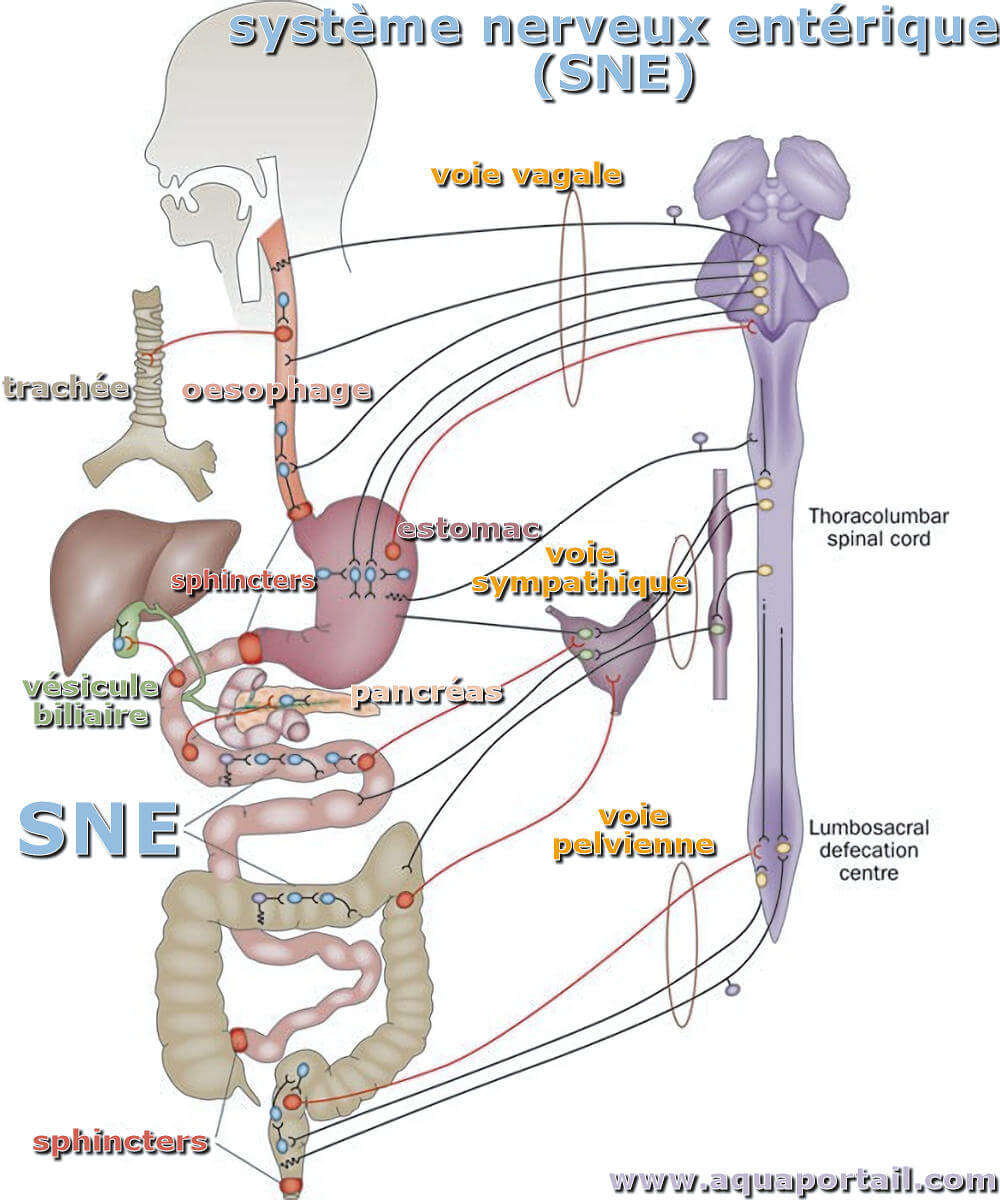
Le système nerveux entérique (SNE) est le système nerveux intrinsèque du tractus gastro-intestinal.
Généralités
Le SNE est une division du système nerveux autonome, les autres divisions étant le sympathique et le parasympathique, avec lesquels il a des connexions étendues.
Rôle et fonctions
Le système nerveux entérique (SNE) contient des circuits réflexes complets qui détectent l'état physiologique du tractus gastro-intestinal, intègrent des informations sur l'état du tractus gastro-intestinal et fournissent des sorties pour contrôler les mouvements de l'intestin, les échanges de fluides entre l'intestin et sa lumière et le flux sanguin local.
C'est la seule partie du système nerveux périphérique (SNP) qui contient des circuits neuronaux étendus capables d'une fonction locale et autonome. Le SNE a de vastes connexions bidirectionnelles avec le système nerveux central (SNC) et travaille de concert avec le SNC pour contrôler le système digestif dans le contexte des demandes physiologiques locales et corporelles.
En raison de son étendue et de son degré d'autonomie, le SNE a été qualifié de deuxième cerveau. Les rôles de le SNE sont beaucoup plus restreints que le cerveau réel, et donc cette analogie a une utilité limitée.
Organisation et relations
Le système nerveux entérique est composé de milliers de petits ganglions situés dans les parois de l'oesophage, de l'estomac, de l'intestin grêle et du gros intestin, du pancréas, de la vésicule biliaire et de l'arbre biliaire, des fibres nerveuses qui relient ces ganglions et des fibres nerveuses qui alimentent le muscle de la paroi intestinale, l'épithélium muqueux, les artérioles et autres tissus effecteurs. Un grand nombre de neurones sont contenus dans le système nerveux entérique, environ 200 à 600 millions chez l'homme. C'est beaucoup plus de neurones que ce qui existe dans tout autre organe périphérique et est similaire au nombre de neurones dans la moelle épinière.
Les ganglions contiennent des neurones et des cellules gliales, mais pas d'éléments du tissu conjonctif, et à bien des égards, leur structure est similaire à celle du SNC, sauf qu'il n'y a pas de barrière hémato-entérique significative du système nerveux. Les faisceaux de fibres nerveuses au sein du système nerveux entérique se composent des axones des neurones entériques, des axones des neurones extrinsèques qui se projettent sur la paroi intestinale et des cellules gliales.
On trouve deux grands ensembles de ganglions, les ganglions mésentériques entre les couches musculaires externes et les ganglions sous-muqueux. Le plexus mésentérique forme un réseau continu, autour de la circonférence de l'intestin et s'étendant de la partie supérieure de l'oesophage au sphincter anal interne. Le plexus sous-muqueux ganglionné est présent dans l'intestin grêle et le gros intestin, mais est absent de l'oesophage et ne contient que très peu de ganglions dans l'estomac.
Le système nerveux entérique provient des cellules de la crête neurale qui colonisent l'intestin pendant la vie intra-utérine. Il devient fonctionnel dans le dernier tiers de la gestation chez l'homme et continue de se développer après la naissance.
Le système nerveux entérique reçoit des apports des parties parasympathiques et sympathiques du système nerveux, et le tractus gastro-intestinal reçoit également un approvisionnement abondant de fibres nerveuses afférentes, à travers les nerfs vagues et les voies afférentes rachidiennes. Ainsi, il existe une riche interaction, dans les deux sens, entre le système nerveux entérique, les ganglions prévertébraux sympathiques et le SNC.
Le tractus gastro-intestinal abrite également un système de signalisation endocrinien étendu, et de nombreuses fonctions gastro-intestinales sont sous double contrôle neuronal et endocrinien. Les neurones entériques interagissent également avec le système immunitaire intrinsèque étendu du tractus gastro-intestinal.
Types de neurones entériques
Environ 20 types de neurones entériques peuvent être définis par leurs fonctions. Des combinaisons de caractéristiques (morphologie, propriétés neurochimiques, physiologie cellulaire et projections vers des cibles) aident à définir chaque type. Parmi les 20 types, trois classes peuvent être identifiées, les neurones afférents primaires intrinsèques (IPAN, également appelés neurones sensoriels intrinsèques), les interneurones et les motoneurones
Les IPAN détectent l'état physique des organes (par exemple, la tension dans la paroi intestinale) et les caractéristiques chimiques du contenu luminal. Ils réagissent à ces signaux pour initier un contrôle réflexe approprié de la motilité, de la sécrétion et du flux sanguin. Les IPAN se connectent entre eux, avec des interneurones et directement avec des motoneurones. Les interneurones se connectent avec d'autres interneurones et avec des motoneurones. Parmi les motoneurones se trouvent les motoneurones musculaires, les neurones sécrétomoteurs, les neurones sécrétomoteurs-vasodilatateurs et les neurones vasodilatateurs.
Contrôle de la motilité
Le tractus gastro-intestinal a une couche musculaire externe dont les buts sont de mélanger les aliments afin qu'ils soient exposés aux enzymes digestives et à la paroi absorbante de l'intestin, et de propulser le contenu du tube digestif. Le muscle se détend également pour s'adapter à une masse accrue de contenu, notamment dans l'estomac.
Chez l'homme en particulier, le côlon a également une fonction de réservoir importante pour retenir les selles jusqu'à la défécation. Les circuits réflexes entériques régulent le mouvement en contrôlant l'activité des neurones excitateurs et inhibiteurs qui innervent le muscle. Ces neurones ont des co-transmetteurs, pour les neurones excitateurs, l'acétylcholine et les tachykinines, et pour les neurones inhibiteurs l'oxyde nitrique, le peptide intestinal vasoactif (VIP) et l'ATP. Il existe également des preuves que le peptide d'activation de l'adénylate cyclase hypophysaire (PACAP) et le monoxyde de carbone (CO) contribuent à la transmission inhibitrice.
Les temps de passage du contenu dans le tractus gastro-intestinal varient en fonction de la nature de l'aliment, y compris sa quantité et sa teneur en nutriments. L'activité péristaltique de l'oesophage transporte les aliments de la bouche à l'estomac en environ 10 secondes, où les aliments sont mélangés aux sucs digestifs. La vidange gastrique se déroule sur des périodes d'environ 1 à 2 heures après un repas, le contenu liquéfié étant propulsé par des ondes péristaltiques gastriques sous forme de petites aspirations dans le jéjunum pendant cette période.
Le liquide de l'estomac est mélangé avec des sécrétions pancréatiques et biliaires pour former le contenu liquide de l'intestin grêle, connu sous le nom de chyme. Le chyme est mélangé et se déplace lentement le long de l'intestin, sous le contrôle de mouvements de mélange et de propulsion orchestrés par l'ENS, tandis que la digestion et l'absorption des nutriments interviennent. Le temps de transit moyen dans l'intestin grêle humain est de 3 à 4 heures. Le transit colique chez les humains en bonne santé prend 1 à 2 jours.
Les réflexes intrinsèques du système nerveux entérique sont essentiels à la génération des schémas de motilité de l'intestin grêle et du gros intestin. Les principaux mouvements musculaires dans l'intestin grêle sont : l'activité de mélange; réflexes propulsifs qui ne parcourent que de petites distances; le complexe myoélectrique en migration; ruées péristaltiques; et la rétropulsion associée aux vomissements. Le système nerveux entérique est programmé pour produire ces différents résultats. Contrairement à l'intestin, le péristaltisme dans l'estomac est une conséquence d'événements électriques conduits (ondes lentes) qui sont générés dans le muscle. L'intensité de la contraction gastrique est déterminée par les actions des nerfs vagues, qui forment des connexions avec les neurones entériques dans les ganglions mésentériques.
L'estomac proximal se détend pour s'adapter à l'arrivée des aliments. Cette relaxation est également médiée par les connexions du nerf vague avec les neurones entériques. Ainsi, les principaux centres d'intégration pour le contrôle de la motilité gastrique se trouvent dans le tronc cérébral, tandis que ceux pour le contrôle de l'intestin grêle et du gros intestin se trouvent dans le système nerveux entérique.
Chez la plupart des mammifères, le tissu contractile de la paroi externe de l'oesophage est un muscle strié, et chez d'autres, y compris l'homme, la moitié proximale ou plus est un muscle strié. La partie musculaire striée de l'oesophage est contrôlée, via le nerf vague, par un circuit d'intégration dans le tronc cérébral. Ainsi, bien que les ganglions mésentériques soient proéminents dans la partie musculaire striée de l'oesophage, ils sont des modificateurs, et non des centres de contrôle essentiels, pour le péristaltisme oesophagien.
Les sphincters des muscles lisses restreignent et régulent le passage du contenu luminal entre les régions. En général, les réflexes initiés en amont des sphincters détendent le muscle du sphincter et facilitent le passage du contenu, tandis que les réflexes initiés de manière distale restreignent le passage rétrograde du contenu dans les parties plus proximales du tube digestif.
La progression du contenu dans une direction orale à anale est inhibée lorsque l'activité nerveuse sympathique augmente. Pour y parvenir, la transmission des réflexes entériques excitateurs vers le muscle est inhibée et les sphincters sont contractés. Les neurones sympathiques post-ganglionnaires utilisent la noradrénaline comme transmetteur principal. Au repos, les voies sympathiques exercent peu d'influence sur la motilité. Ils entrent en action lorsque les réflexes protecteurs sont activés.
Echanges de fluides et du flux sanguin local
Le système nerveux entérique régule le mouvement de l'eau et des électrolytes entre la lumière intestinale et les compartiments des fluides tissulaires. Pour ce faire, il dirige l'activité des neurones sécrétomoteurs qui innervent la muqueuse de l'intestin grêle et du gros intestin et contrôlent sa perméabilité aux ions. Les neurotransmetteurs des neurones sécrétomoteurs sont le peptide intestinal vasoactif (VIP) et l'acétylcholine. La sécrétion est intégrée à la vasodilatation, qui fournit une partie du liquide sécrété. La plupart des neurones sécrétomoteurs ont des corps cellulaires dans les ganglions sous-muqueux.
Des flux de liquide, supérieurs au volume sanguin total du corps, traversent chaque jour les surfaces épithéliales du tractus gastro-intestinal. Le contrôle de ce mouvement de fluide via le système nerveux entérique est d'une importance primordiale pour le maintien de l'équilibre hydrique et électrolytique de l'ensemble du corps. Les flux les plus importants se font à travers l'épithélium de l'intestin grêle, avec un mouvement de fluide important intervenant également dans le gros intestin, l'estomac, le pancréas et la vésicule biliaire. L'eau se déplace entre les lumières des organes digestifs et les compartiments des fluides corporels en réponse au transfert de molécules osmotiquement actives. La plus grande absorption d'eau, 8–9 litres par jour, accompagne les flux entrants de molécules nutritives et de Na+ par l'activation de co-transporteurs de nutriments, et la plus grande sécrétion accompagne les flux sortants de Cl- et HCO3- dans l'intestin grêle et le gros intestin, vésicule biliaire et pancréas. Dans chacun de ces organes, la sécrétion liquidienne est contrôlée par des réflexes entériques. Dans l'intestin grêle et la majeure partie du côlon, les circuits réflexes sont intrinsèques, dans le système nerveux entérique. Ils équilibrent la sécrétion avec des flux absorbants, et puisent l'eau du fluide absorbé et de la circulation. L'activité des réflexes sécrétomoteurs est sous un contrôle physiologiquement important des voies nerveuses sympathiques inhibitrices qui répondent aux changements de pression artérielle et de volume sanguin par les centres réflexes centraux.
Le flux sanguin local vers la muqueuse est régulé par les neurones vasodilatateurs entériques de sorte que le flux sanguin muqueux est approprié pour équilibrer les besoins nutritifs de la muqueuse et pour accueillir l'échange de fluide entre le système vasculaire, le liquide interstitiel et la lumière intestinale. Il n'y a pas de neurones vasoconstricteurs intrinsèques. Le flux sanguin global vers l'intestin est régulé à partir du SNC, via des neurones vasoconstricteurs sympathiques. Les neurones vasoconstricteurs sympathiques agissent de concert avec le contrôle autonome d'autres lits vasculaires, pour distribuer le débit cardiaque en fonction des besoins relatifs de tous les organes. Ainsi, en cas de besoin, même pendant la digestion, le sympathique peut détourner le flux sanguin du tractus gastro-intestinal.
Sécrétions gastriques et pancréatiques
La sécrétion d'acide gastrique est régulée à la fois par les neurones et par les hormones. La régulation neurale se fait par des neurones cholinergiques avec des corps cellulaires dans la paroi de l'estomac. Ceux-ci reçoivent des entrées excitatrices à la fois de sources entériques et des nerfs vagues.
La sécrétion gastrique de HCl et de pepsinogène dans l'estomac et la sécrétion d'enzymes pancréatiques dépendent en grande partie des réflexes vago-vagaux. Les motoneurones entériques sont la dernière voie commune, mais les rôles des réflexes intrinsèques sont mineurs. La sécrétion pancréatique de bicarbonate, pour neutraliser le contenu duodénal, est contrôlée par la sécrétine, une hormone libérée par le duodénum, en synergie avec l'activité des neurones entériques cholinergiques et non cholinergiques. La sécrétion dans la vésicule biliaire et la sécrétion de bicarbonate dans l'estomac distal sont également contrôlées par les nerfs.
Cellules endocrines gastro-intestinales
Les fibres nerveuses sont proches des cellules endocrines de la muqueuse du tractus gastro-intestinal, dont certaines sont sous contrôle neural. Par exemple, les cellules de gastrine dans l'antre de l'estomac sont innervées par des neurones excitateurs qui utilisent le peptide libérant de la gastrine comme neurotransmetteur principal. A l'inverse, les hormones libérées par les cellules endocrines gastro-intestinales influencent les terminaisons des neurones entériques. Dans un sens, les cellules endocrines agissent comme des cellules gustatives, qui échantillonnent l'environnement luminal et libèrent des molécules messagères dans le tissu de la muqueuse, où se trouvent les terminaisons nerveuses. C'est une communication nécessaire, car les terminaisons nerveuses sont séparées de la lumière par l'épithélium muqueux. Une communication importante est avec la sérotonine (5-hydroxytryptamine, 5-HT) contenant des cellules endocrines qui activent les réflexes de motilité. Une libération excessive de sérotonine peut provoquer des nausées et des vomissements, et les antagonistes des récepteurs 5-HT3 sont des anti-nauséeux.
Réactions de défense
Les neurones entériques sont impliqués dans un certain nombre de réactions de défense de l'intestin. Les réactions de défense comprennent la diarrhée pour diluer et éliminer les toxines, une activité propulsive colique exagérée qui intervient lorsqu'il y a des agents pathogènes dans l'intestin et des vomissements.
La sécrétion liquidienne est provoquée par des stimuli nocifs, notamment par la présence intraluminale de certains virus, bactéries et toxines bactériennes. Cette sécrétion est due en grande partie à la stimulation des réflexes sécrétomoteurs entériques. Le but physiologique est sans aucun doute de débarrasser l'organisme des agents pathogènes et de leurs produits. Cependant, si les agents pathogènes dépassent la capacité du corps à faire face, la perte de liquide (diarrhée) peut devenir une menace sérieuse pour l'organisme.
Réflexes entéro-entériques
Les signaux entre les régions intestinales sont transportés à la fois par des hormones (telles que la cholécystokinine, la gastrine et la sécrétine) et par des circuits nerveux. Les réflexes entéro-entériques régulent une région par rapport aux autres. Par exemple, lorsque les nutriments pénètrent dans l'intestin grêle, la sécrétion d'enzymes digestives par le pancréas intervient. Une série de circuits nerveux qui transportent des signaux d'une région de l'intestin vers les ganglions sympathiques et retournant à la paroi intestinale fournissent un système de régulation unique au tractus gastro-intestinal. Les neurones avec des corps cellulaires dans les ganglions entériques et des terminaisons dans les ganglions sympathiques prévertébraux forment les membres afférents de ces réflexes. Ceux-ci sont connus sous le nom de neurones afférents intestinofugaux (IFAN).
Interactions SNE-SNC
Le tractus gastro-intestinal est en communication bidirectionnelle avec le SNC (système nerveux central). Les neurones afférents transmettent des informations sur l'état du tractus gastro-intestinal. Une partie de cela atteint la conscience, y compris la douleur et l'inconfort de l'intestin et les sentiments conscients de faim et de satiété, qui sont des perceptions intégrées dérivées du tractus gastro-intestinal et d'autres signaux (glycémie, par exemple). D'autres signaux afférents, concernant par exemple la charge nutritive dans l'intestin grêle, ou l'acidité de l'estomac, n'atteignent normalement pas la conscience. À son tour, le SNC fournit des signaux pour contrôler l'intestin, qui sont, dans la plupart des cas, relayés par le SNE.
Par exemple, la vue et l'odeur des aliments provoquent des événements préparatoires dans le tractus gastro-intestinal, notamment la salivation et la sécrétion d'acide gastrique. C'est ce qu'on appelle la phase céphalique de la digestion. Les aliments avalés stimulent le pharynx et la partie supérieure de l'oesophage, provoquant des signaux afférents qui sont intégrés dans le tronc cérébral, puis fournissent des signaux efférents aux neurones entériques de l'estomac qui provoquent une sécrétion acide et une augmentation du volume gastrique, en vue de l'arrivée de la nourriture. À l'autre extrémité de l'intestin, les signaux du côlon et du rectum sont transmis aux centres de défécation de la moelle épinière, à partir desquels un ensemble programmé de signaux est transmis au côlon, au rectum et au sphincter anal pour provoquer la défécation.
Les centres de défécation sont sous contrôle inhibiteur des régions supérieures du SNC, et une inhibition qui peut être libérée lorsqu'il est choisi de déféquer. Les autres influences centrales passent par les voies sympathiques, qui ont été discutées dans les sections sur le contrôle de la motilité et la régulation des échanges de fluides et du flux sanguin local, ci-dessus.
En rapport avec "système nerveux entérique"
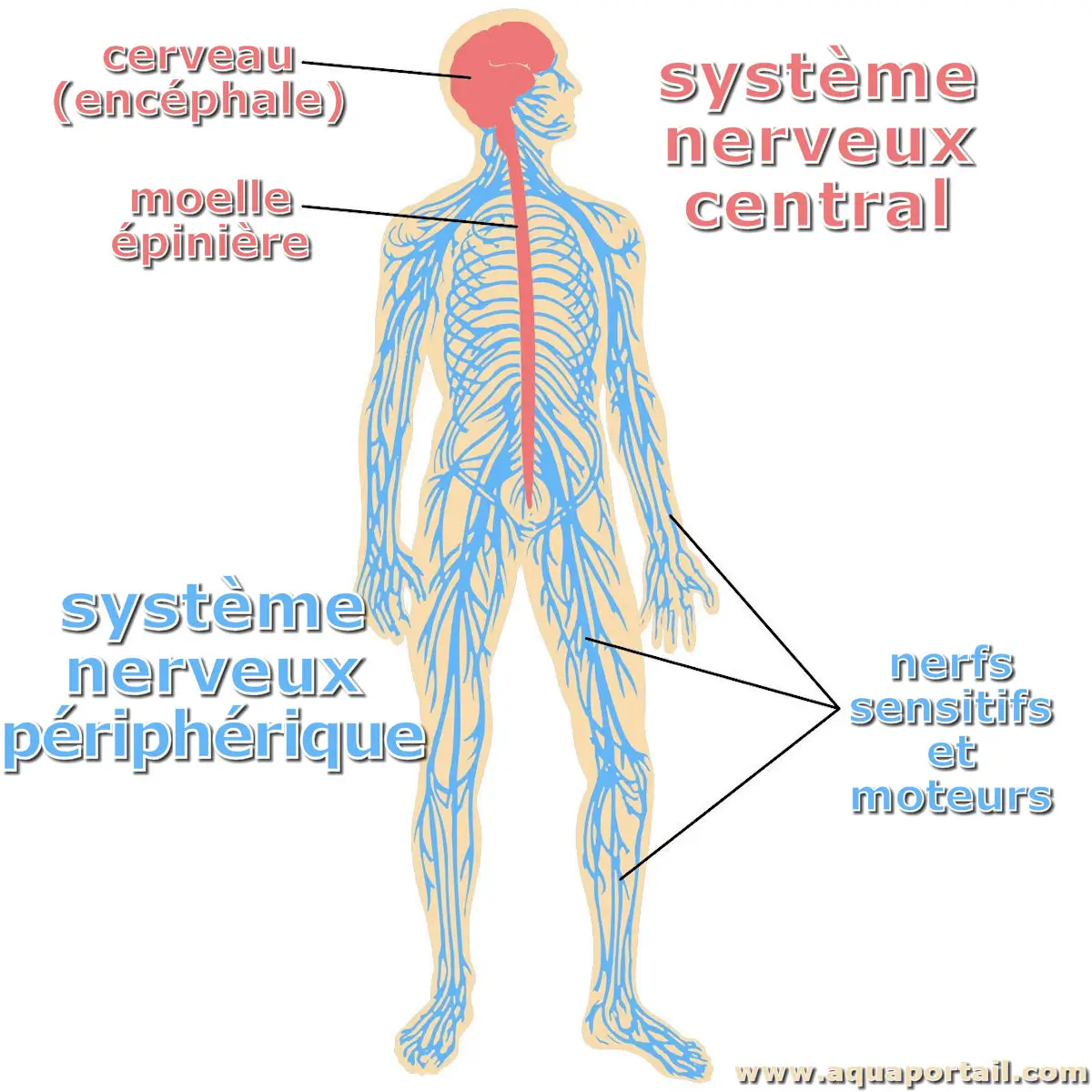
Le système nerveux d'un organisme est un réseau complexe composé de l'encéphale et de la moelle épinière (système nerveux central), et des nerfs (système...
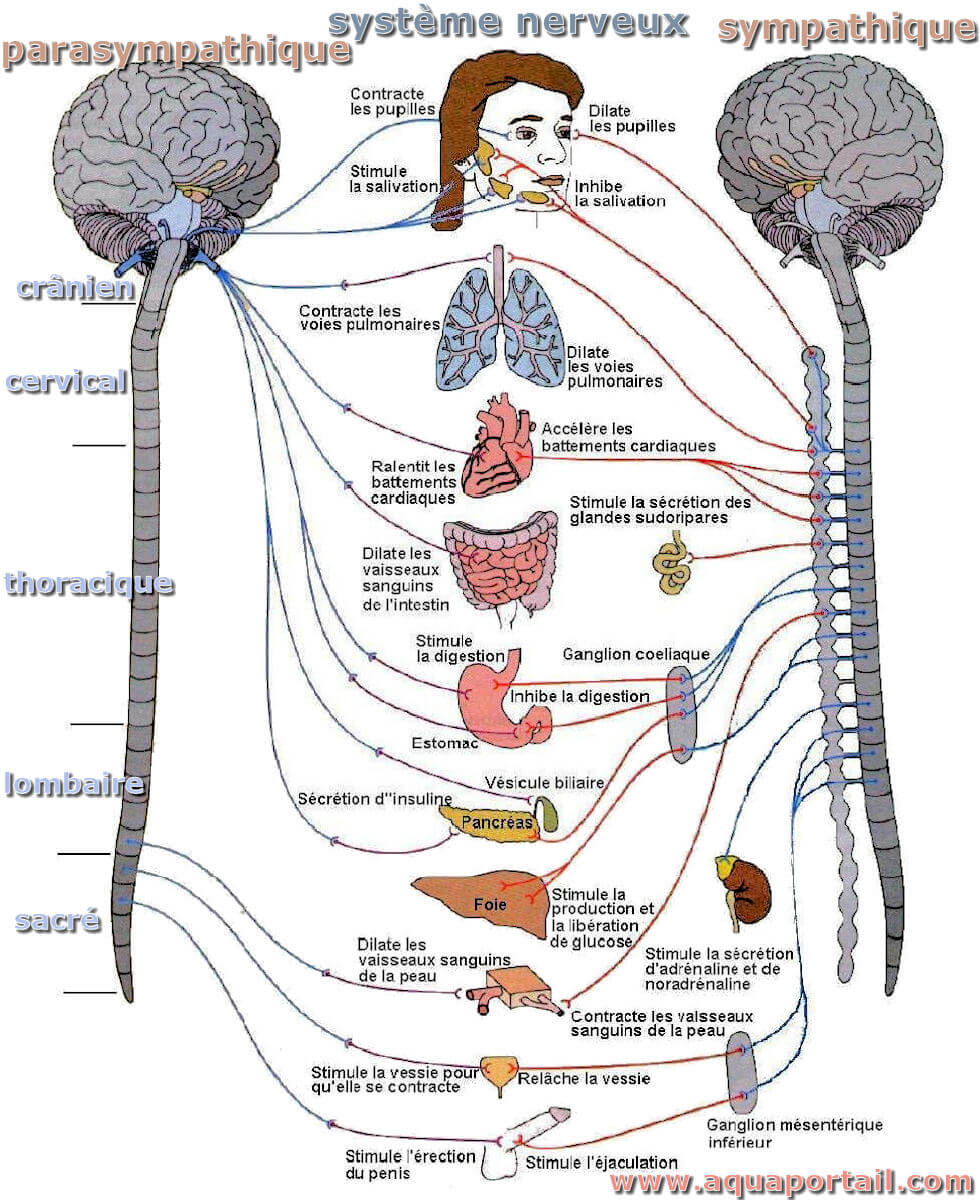
Le système nerveux autonome régule les processus physiologiques involontaires, notamment la fréquence cardiaque, la tension artérielle, la respiration...
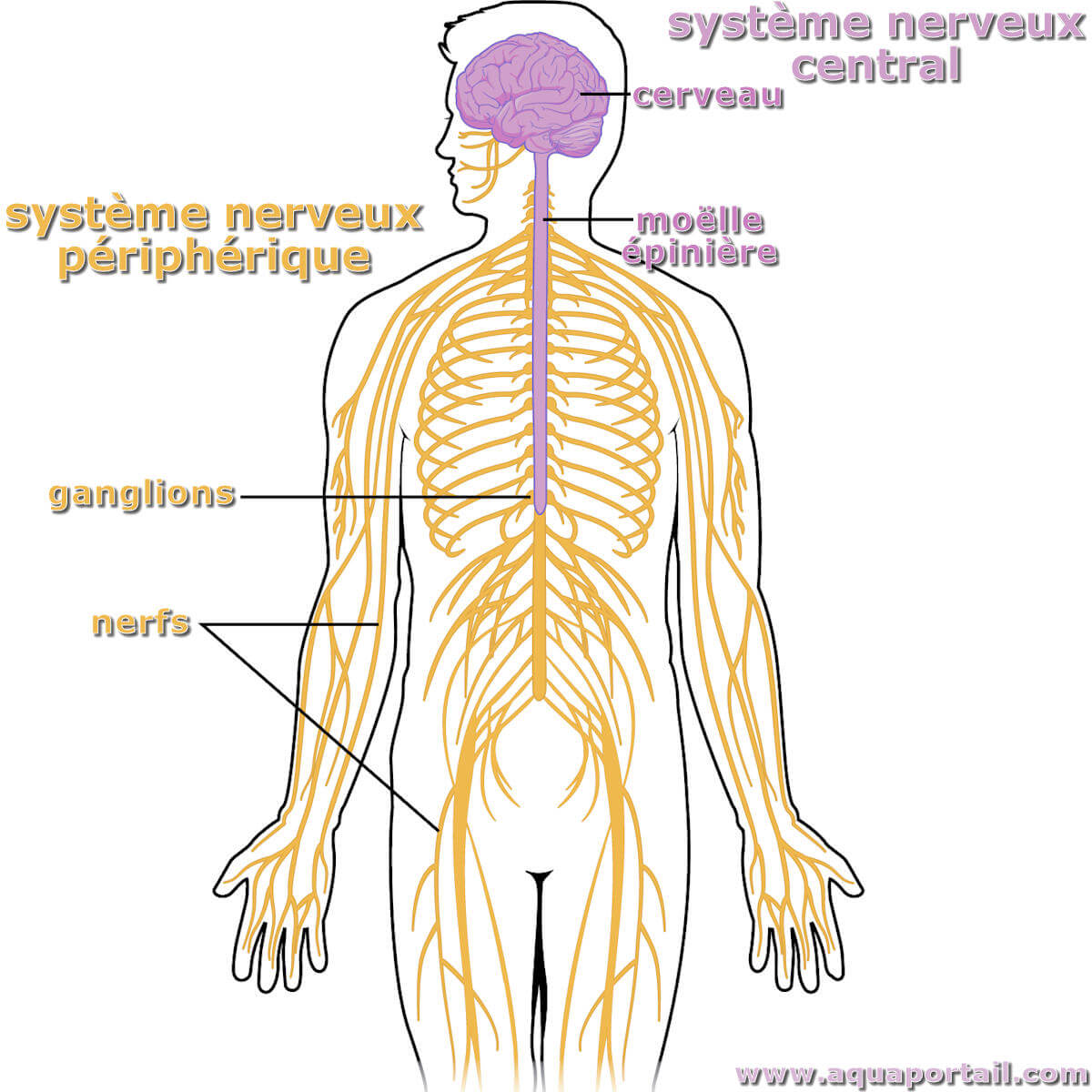
Le système nerveux central (SNC) est une division anatomique du système nerveux qui est constituée des tissus nerveux du cerveau et de la moelle épinière.
système nerveux parasympathique
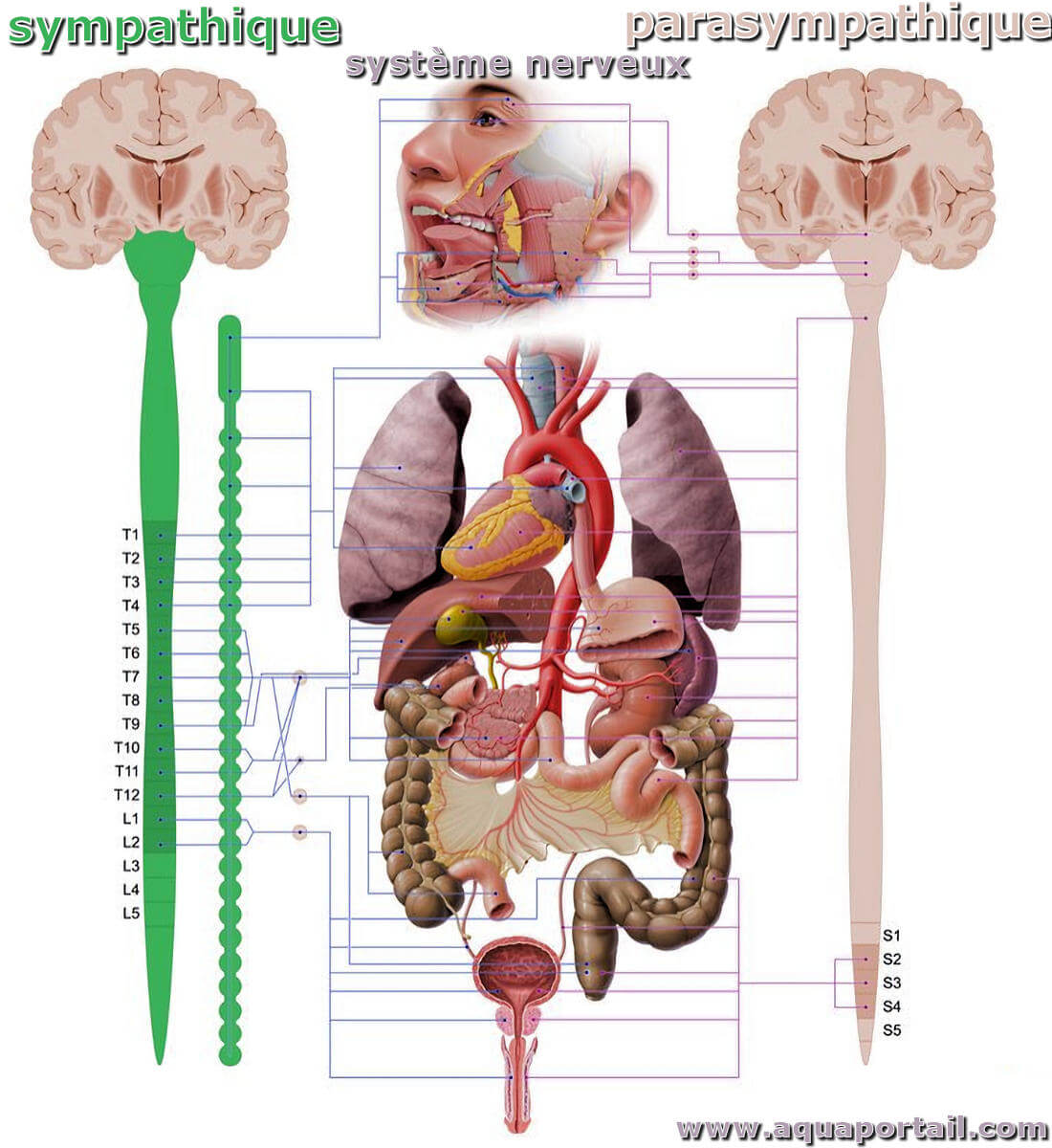
Le système nerveux parasympathique est une division du système nerveux autonome (SNA) qui contrôle l'activité des muscles lisses et cardiaques et des glandes.