
Polyspermie
Définition
La polyspermie est la fécondation d'un gamète femelle par plusieurs gamètes mâles. Plusieurs spermatozoïdes pénètrent dans l'ovule dans un événement de fécondation polyspermique.
La polypsermie est bloquée par la zone pellucide :
Parmi les blocages à la polyspermie, la zone pellucide joue une rôle majeur.
Explications
La polyspermie intervient normalement dans les ovules avec beaucoup de jaune vitellin (comme chez les oiseaux, les requins et les chimères), mais alors seulement un spermatozoïde fusionne avec le noyau de l'ovule.
Les mécanismes du phénomène de la polyspermie sont encore inconnus.
Effets de la polyspermie
Bien qu'il y ait beaucoup de spermatozoïdes qui peuvent se lier à un ovule, normalement un seul fusionne avec la membrane plasmique de l'ovule et injecte son noyau et d'autres organites dans le cytoplasme de l'ovule. Si la fusion de plus d'un spermatozoïde existe, et donc le phénomène de polyspermie intervient, la formation mitotique d'ensembles mitotiques (fuseaux mitotiques) supplémentaires ou multipolaires est générée, ce qui provoque la ségrégation défectueuse des chromosomes au cours de la division cellulaire; des cellules non diploïdes sont formées et le développement s'arrête généralement.
Chez l'oursin, la fécondation par deux spermatozoïdes engendre un noyau triploïde, dans lequel chaque chromosome est représenté trois fois au lieu de deux. Pire encore, parce que le centriole fourni par chaque spermatozoïde se divise pour former les deux pôles de l'appareil mitotique, les chromosomes triploïdes seront déplacés de quatre positions différentes par des fils de broche, formant quatre cellules qui auront une charge chromosomique différente. Ces cellules meurent ou se développent anormalement.
Chez l'homme, la polyploïdie (fécondation multiple) est généralement mortelle pour l'embryon et a été détectée dans les cas d'avortement spontané d'environ 10 à 20 %, notamment an cas de dispermie.
Puis, en raison de l'infaisabilité de l'embryon après un événement de polyspermie, et malgré l'union de nombreux spermatozoïdes à la couche entourant l'ovule, il est important qu'un seul spermatozoïde fusionne avec la membrane plasmique et délivre son noyau dans cette zone. Pour cela, les animaux ont développé deux mécanismes principaux.
Blocage de la polyspermie
En raison de l'inéligibilité de l'embryon après un événement de polyspermie et malgré l'union de nombreux spermatozoïdes à la couche entourant l'ovule, il est important qu'un seul spermatozoïde ne fusionne avec la membrane plasmatique et en délivre le noyau. Pour cela, les animaux ont développé deux mécanismes principaux.
Blocage rapide
Le blocage rapide de la polyspermie est obtenu par une modification du potentiel électrique de la membrane plasmatique (ou oeuf) de l'oeuf, qui agit comme une barrière sélective entre le cytoplasme et l'environnement extracellulaire. Ainsi, la concentration ionique diffère significativement d'un côté à l'autre, notamment au niveau des ions sodium (Na+) et du potassium (K+). Dans l'eau de mer, la concentration externe de sodium est particulièrement élevée. C'est pour cette raison que ce mécanisme est particulièrement utilisé par les invertébrés marins.
Ainsi, ce type de blocage est réalisé par l'ouverture de canaux dépendant du sodium qui permettent à l'ion de pénétrer dans le cytoplasme cellulaire, en dépolarisant la membrane, c'est-à-dire en augmentant les charges positives à l'intérieur (généralement le potentiel de la cellule). la membrane, appelée potentiel de repos, se situe à -70 mV, ce qui indique que les charges négatives sont plus grandes dans la cellule, avec la fusion du sperme, le potentiel atteint des valeurs positives).
Une expérience dans laquelle l'importance du sodium dans le blocage rapide de la polyspermie a été démontrée par Laurinda Jaffe et ses collègues, modifiant la concentration de cet ion dans l'environnement. Ainsi, la réduction de la concentration en sodium a augmenté le nombre d'oeufs polyspermiques, car l'ouverture des canaux n'a pas entraîné l'entrée de suffisamment de molécules dans le cytoplasme et le changement nécessaire de potentiel n'a donc pas été observé.
Il n'y a aucune certitude quant à la manière dont une modification du potentiel membranaire agit sur le sperme pour bloquer la fertilisation secondaire. L'hypothèse la plus acceptée est que le spermatozoïde charge un composant sensible à la tension (peut-être une protéine fusogène pour la fusion des membranes chargées positivement) dont l'insertion dans le cytoplasme dépend de la charge électrique le long de la membrane de l'oeuf.
Chez Rana pipiens, la grenouille léopard, le potentiel de la membrane passe à +8 mV, une seconde; augmentant lentement jusqu'à +17 mV. Ce potentiel est maintenu pendant 20 minutes. Cependant, ce mécanisme n'intervient pas chez les salamandres, et il n'est pas non plus utile d'éviter la polyspermie dans la fécondation croisée des oeufs de xénope Xenopus tropicalis par le sperme de la salamandre. Un blocage rapide par dépolarisation membranaire a également été démontré chez le ver marin Urechis caupo, qui a également testé l'importance du changement de potentiel en soi plus que le flux d'ions dans deux cas d'espèces d'algues fucoïdes (des fucus, une famille d'algues brunes) et d'étoiles de mer.
Alors que les espèces non mammifères utilisent ce type de blocage membranaire, chez les mammifères, il agit par un mécanisme différent, car aucune dépolarisation membranaire n'est observée dans les oeufs fécondés de souris, de hamster et de lapin.
Blocage lent
Un second mécanisme pour empêcher l'entrée de plusieurs spermatozoïdes dans le cytoplasme de l'ovule est nécessaire car le mécanisme rapide est transitoire : le potentiel membranaire d'un ovule reste positif pendant une courte période seulement. Ainsi, le second mécanisme implique un événement de longue durée, la réaction dite des granules corticaux.
Lorsque le sperme fusionne avec la membrane plasmique de l'ovule, il génère une augmentation locale du calcium cytosolique, qui se disperse à travers la cellule dans une onde. Il existe des preuves à l'appui de l'idée que l'onde ou les oscillations (chez certains mammifères présentant des oscillations plutôt que des ondes) du Ca2+ sont induites par une protéine introduite par les spermatozoïdes. Cependant, chez les oursins et les mammifères, l'augmentation de la concentration en calcium n'est pas due à l'entrée de cet ion dans le milieu extracellulaire, mais plutôt à la libération de l'élément des dépôts intracellulaires. Ainsi, les ions calcium sont stockés dans le réticulum endoplasmique de l'ovule et sont libérés par l'activation des canaux calciques dans la membrane du réseau dépendant de l'inositol triphosphate (IP3). Ce composé est activé précisément par contact avec des protéines dans le sperme. Chez les oursins et les grenouilles, le réticulum endoplasmique est prononcé dans le cortex et entoure les granules corticaux, le calcium étant directement responsable de la propagation de la réaction corticale. Une fois expulsés du dépôt, les ions libres ont la capacité d'induire l'expulsion du calcium encore séquestré dans les sites de stockage, générant ainsi l'onde de propagation et l'exocytose des granules corticaux.
Le mécanisme de l'exocytose est le suivant. Directement sous la membrane plasmique de l'ovule se trouvent des milliers de granules corticaux (environ 15 000 chez le hérisson), activés pour réaliser l'exocytose par l'onde de calcium propagée juste après l'entrée du sperme. Grâce à ce processus, les vésicules situées dans le cytoplasme fusionnent avec la membrane et libèrent leur contenu. Chez le hérisson, les granules libèrent plusieurs protéines dans l'espace entre la membrane cytoplasmique de l'oeuf et le tapis fibreux des protéines dans l'enveloppe vitelline.
Parmi les protéines libérées figurent des protéases qui dégradent les jonctions de connexion entre l'enveloppe de vitelline et la membrane cellulaire, en plus d'éliminer les récepteurs de liaison (ceux auxquels se lient les spermatozoïdes) et de tous les spermatozoïdes attachés à l'oeuf. De plus, des mucopolysaccharides sont libérés, générant un gradient osmotique et entraînant la pénétration de l'eau dans cet espace et l'expansion de l'enveloppe, générant la membrane de fertilisation. Une troisième enzyme libérée, une peroxydase, renforce la membrane de fertilisation en réticulant les résidus de tyrosine dans les protéines adjacentes.
La formation de la membrane de fertilisation commence au point d'entrée du sperme et continue son expansion à travers le reste de l'oeuf. Ce procédé donne également la formation de la couche hyaline, par la libération d'une quatrième protéine portant ce nom, mais dont la fonction est de supporter les blastomères lors du clivage.
Blocage chez les poissons
Contrairement à de nombreux autres organismes, le contact entre le sperme et l'oeuf d'un poisson n'existe qu'une fois que le gamète mâle est entré par un canal, la micropile, à un site spécifique de l'oeuf. Peut-être pour cette raison la plupart des poissons n'ont pas d'acrosome dans leurs spermatozoïdes.
Par conséquent, les mécanismes de blocage de la polyspermie chez les poissons diffèrent considérablement. Ce processus est effectué en au moins quatre étapes. Tout d'abord, il y a des changements dans la structure du micropyle, qui se ferme pour réduire le nombre de spermatozoïdes atteignant la membrane plasmique de l'oeuf.
Ensuite, la formation du cône de fécondation, issu de la fusion entre les membranes des deux gamètes, bloque l'ouverture du micropyle. Par la suite, il existe des interactions entre les lectines libérées des granules corticaux et les spermatozoïdes surnuméraires dans le vestibule du micropyle : les protéines immobilisent initialement le sperme et sont ensuite expulsées du vestibule en augmentant la pression osmotique du liquide périvitininique. Agir en seguid, le fluide périvitininique avec les lectines interagit avec l'extérieur de la couche fibreuse, pour éliminer l'orientation et l'attraction du sperme vers le micropyle. Enfin, la réaction de zone intervient, durcissant le chorion et atteignant ainsi la fermeture totale du micropyle.
Blocage chez les mammifères
On pense que plusieurs processus impliquant l'appareil reproducteur de la femelle, tels que l'entraînement et le contact avec le réservoir oviductal, servent à réguler le nombre de spermatozoïdes atteignant le site de fertilisation chez de nombreuses espèces.
Chez les mammifères, le blocage de la polyspermie dans l'ovule existe principalement à deux niveaux : dans la zone pellucide (analogue à l'enveloppe vitellinique chez les non-mammifères) et dans la membrane cellulaire. Il existe donc une différenciation spatiale, mais, contrairement aux espèces non mammifères dans lesquelles la différenciation est également temporaire, chez les organismes de cette classe, il n'y a pas de différence temporelle entre les deux types de blocus.
Le blocage dans la zone pellucide est assez similaire à celui des organismes non mammifères. La fusion des spermatozoïdes avec l'oeuf induit également un signal Ca2+, initié du côté opposé au site de liaison du sperme, qui active l'oeuf pour effectuer la réaction corticale. Cependant, chez les mammifères, la membrane de fertilisation n'est pas créée. Au lieu de cela, la modification des récepteurs dans la zone pellucide intervient, modifiant la composition protéique de la famille. Ce processus est appelé "réaction de zone".
Sans augmentation de la concentration en Ca2+, la libération de granules corticaux ne permet pas de prévenir la reprise de la polyspermie ou de la méiose et l'entrée subséquente dans les cycles cellulaires embryologiques. Des occupations de calcium ont été observées chez les Médaka (poissons), les oursins, les grenouilles, les souris, les hamsters, les bovins, les lapins et les humains.
En revanche, le bloc membranaire des ovules de mammifères diffère largement du mécanisme de dépolarisation retrouvé chez les organismes extérieurs à cette classe. Chez les mammifères, ce bloc semble être l'aboutissement de multiples événements qui, ensemble, modifient la réceptivité au sperme de la membrane de l'ovule.
À l'appui d'un mécanisme multiple, McAvey et ses collaborateurs ont montré que le taux de spermatozoïdes fusionnés par la zone sans oeuf pellucide augmentait lorsqu'ils étaient traités avec un médicament perturbant l'actine dans le cytosquelette, ce qui nécessite certaines fonctions du cytosquelette de la membrane plasmique de l'ovule, et cela n'a aucune implication dans les oscillations de la concentration en calcium ou dans l'exocytose des granules corticaux, similaire à celle des ovules témoins.
De plus, des expériences utilisant des marqueurs fluorescents pour suivre certains lipides membranaires dans des ovules de souris suggèrent qu'il y a des changements dans l'organisation de ceux-ci, induits par la fertilisation. Cependant, nous ne savons pas beaucoup plus que cela. Enfin, il a été montré que les vagues d'augmentation du calcium régulées par l'interaction avec les spermatozoïdes déterminent le temps d'établissement de la membrane bloquante, ce processus étant plus lent dans les ovules présentant une ou plusieurs ondes calciques. Cependant, le calcium ne semble être qu'un facilitateur dans le blocage de la membrane, qui fonctionne avec des événements associés à la fécondation, encore inconnus.
Synonymes, antonymes
Voir tous les synonymes pour "polyspermie".2 synonymes (sens proche) de "polyspermie" :
0 antonyme (sens contraire).
Les mots ou les expressions apparentés à POLYSPERMIE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
Le mot POLYSPERMIE est dans la page 7 des mots en P du lexique du dictionnaire.
Mots en P à proximité
polypode polysaccharide polysépale polysome polysorbate polyspermiepolystique polyténie polytome polytrophique polytypique
En rapport avec "polyspermie"
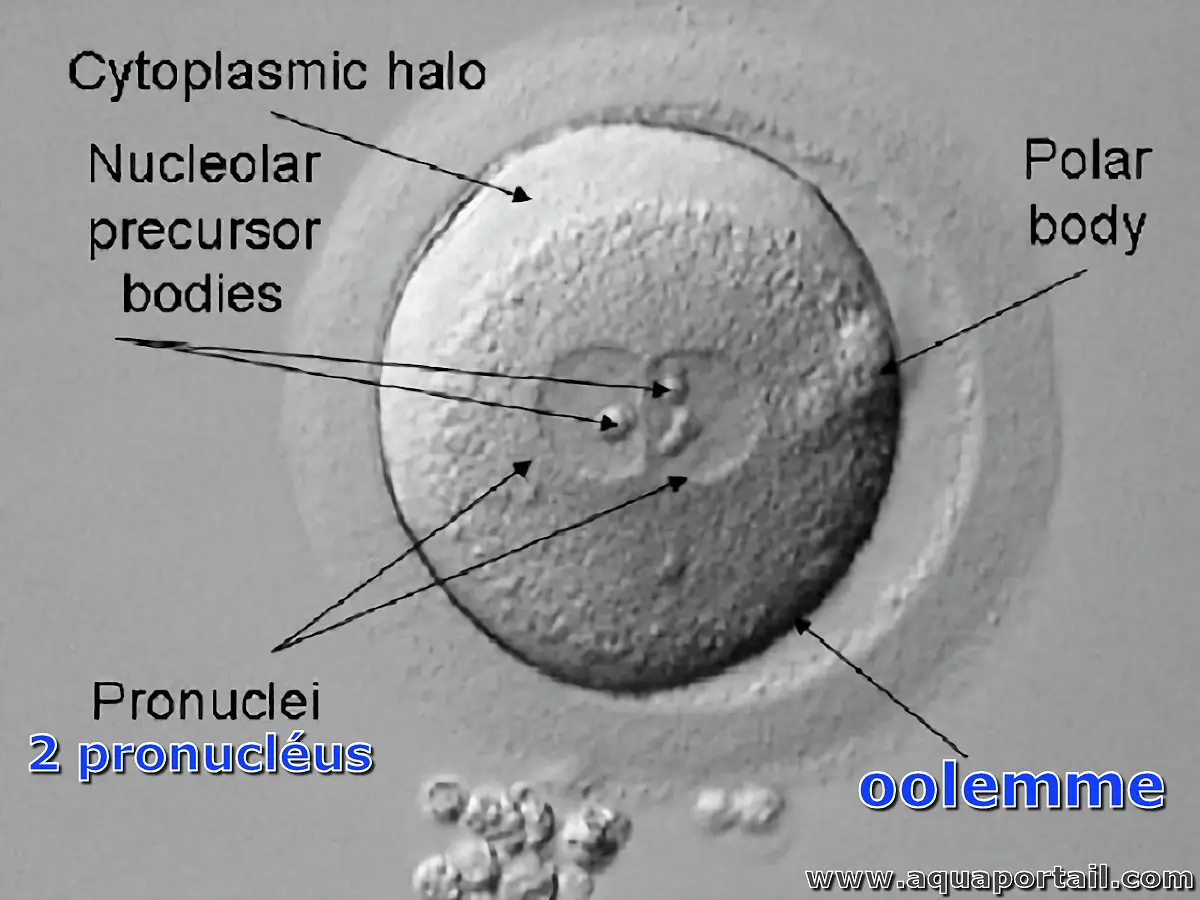
L'oolemme est l'enveloppe externe de la membrane plasmique des ovocytes qui permet de bloquer la polyspermie.

La zone pellucide est une enveloppe extracellulaire relativement épaisse qui entoure tous les ovocytes, les ovules ovulés et les embryons préimplantatoires...

Une agamospermie (apomixie botanique) est le processus par lequel certaines espèces de plantes produisent des graines de manière asexuée, sans fertilisation...
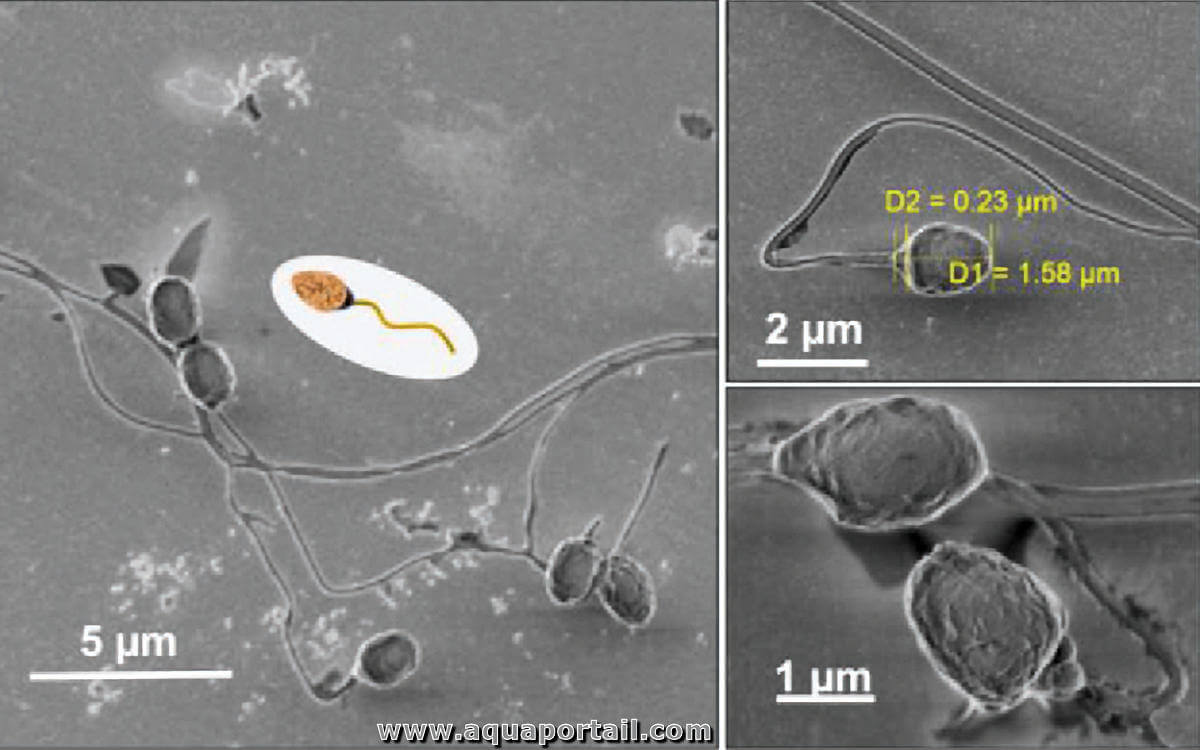
L'aquasperme est un sperme rejeté dans un milieu aquatique tel que l'eau de mer par certains poissons ovulipares et mollusques.