
Membrane mitochondriale interne
Définition
La membrane mitochondriale interne est la barrière fonctionnelle au passage des petites molécules d'une mitochondrie entre le cytosol et la matrice mitochondriale. Elle maintient le gradient de protons qui entraîne la phosphorylation oxydative.
La membrane interne est complétée par une membrane mitochondriale externe qui entoure l'extérieur de la mitochondrie dans une cellule eucaryote. La membrane mitochondriale interne contient d'abondantes protéines de transport responsables de l'établissement d'un gradient de protons entre l'espace intermembranaire et la matrice.
Les membranes mitochondriales :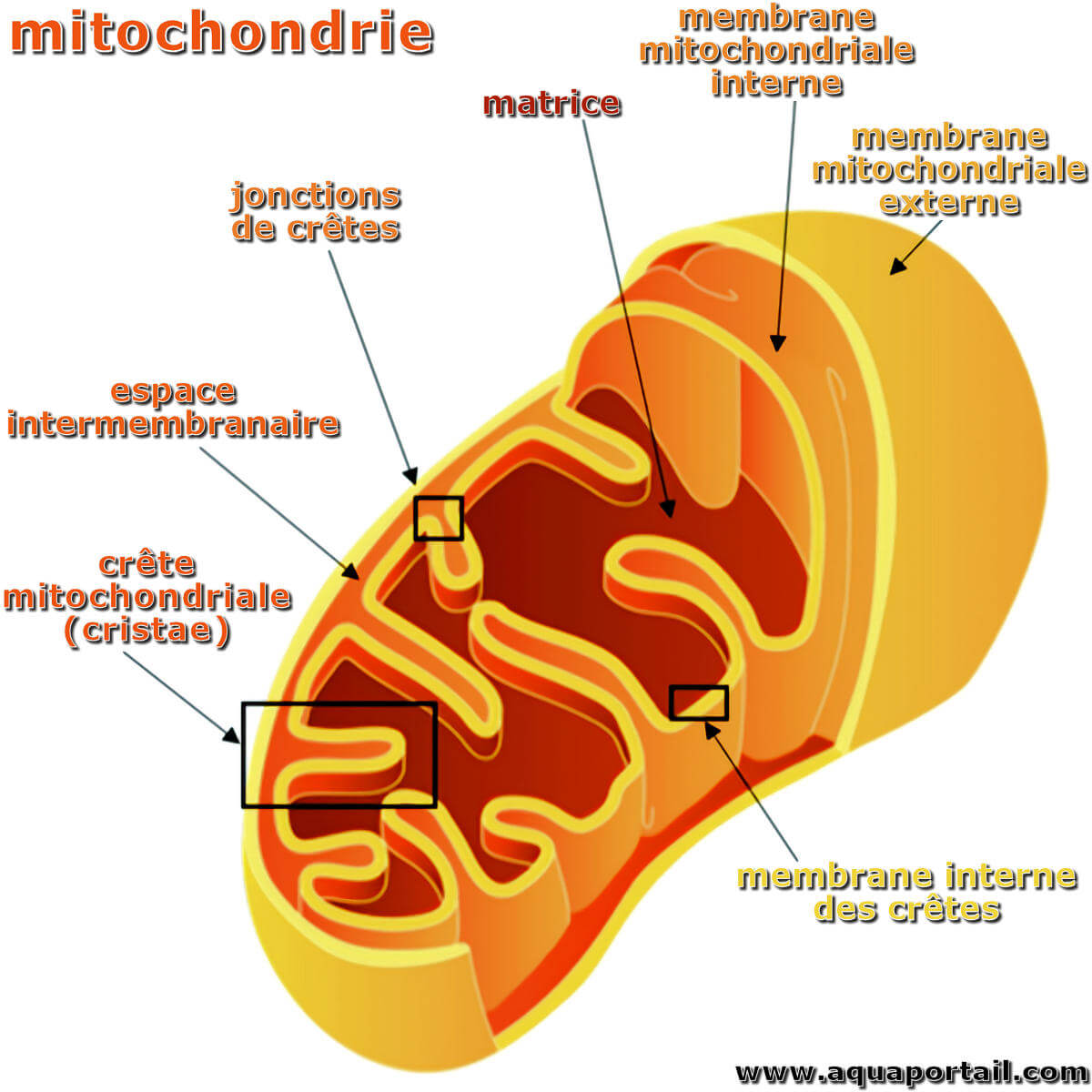
Représentation schématique de l'architecture mitochondriale avec : la membrane mitochondriale externe (MME), la membrane mitochondriale interne (MMI), la membrane interne des crêtes, les jonctions de crêtes (CJ), l'espace intermembranaire, les crêtes mitochondriales (cristae) et la matrice mitochondriale.
Explications
Selon la théorie de l'endosymbiose, la membrane interne des mitochondries aurait été initialement la membrane unique du procaryote ingéré pour générer la double membrane d'une mitochondrie.
Les mitochondries sont entourées de deux membranes phospholipidiques, la membrane mitochondriale externe (MME) et la membrane mitochondriale interne (MMI), qui divisent l'organite en deux espaces, la matrice mitochondriale et l'espace intermembranaire. Les deux membranes présentent des différences significatives dans la composition lipidique, les caractéristiques et les rôles des protéines transmembranaires, la perméabilité et la forme et sont le résultat de l'origine endosymbiotique de l'organite.
La membrane mitochondriale interne héberge les enzymes de la chaîne respiratoire de transport d'électrons, notamment cinq complexes enzymatiques multiprotéiques (I-V) et deux transporteurs d'électrons - la coenzyme Q10 et le cytochrome c. Les cinq complexes contiennent NADH :ubiquinone oxydoréductase (complexe I), succinate :ubiquinone oxydoréductase (complexe II, SQR), ubiquinone :cytochrome c oxydoréductase (complexe III, cytochrome réductase, cytochrome bc1), cytochrome c oxydase (complexe IV, cytochrome oxydase), et ATP synthase (complexe V, F1F0-ATPase).
Importance des deux membranes mitochondriales
En effet, la membrane mitochondriale externe (MME) est plus similaire dans sa composition lipidique aux membranes cellulaires eucaryotes, tandis que la membrane mitochondriale interne (MMI) ressemble aux membranes bactériennes contenant de la cardiolipine. La MMI se caractérise par un rapport protéines/lipides plus élevé et forme des invaginations fortement tassées dans la matrice, appelées crêtes.
Intégrée dans les crêtes réside, avec de nombreuses autres protéines, la machinerie de phosphorylation oxydative (OXPHOS) et l'une des raisons proposées pour le repliement de la MMI est d'augmenter la surface disponible pour la production d'énergie. La partie de la MMI qui ne dépasse pas dans la matrice mais qui, à la place, est parallèle à la MME est appelée la membrane limite interne (IBM). Les cristae et l'IBM sont reliés par des structures tubulaires étroites ou en forme de fente, les jonctions de crêtes (CJ).
De plus, les deux membranes mitochondriales, externe et interne, diffèrent largement par leur perméabilité. Alors que la MME permet le passage d'ions et de petites molécules à travers les canaux anioniques dépendants de la tension (VDAC), seuls l'eau, l'oxygène (O2) et le dioxyde de carbone (CO2) peuvent passer librement à travers la MMI. Cette sélectivité permet la formation d'un gradient électrochimique à travers la membrane, qui constitue la base de la production d'ATP, et la régulation stricte des autres concentrations d'ions, comme le calcium, largement utilisé dans la signalisation cellulaire.
Machinerie de phosphorylation oxydative (OXPHOS)
La membrane mitochondriale interne est le foyer des protéines d'OXPHOS (phosphorylation oxydative) et, à l'intérieur de la membrane, les protéines sont très étroitement regroupées. Les protéines constituent près de 80 % de la masse de la membrane mitochondriale interne, ce qui a des implications importantes pour les mécanismes par lesquels les spécialisations mitochondriales évolutives et développementales surviennent. Par exemple, si un poisson pouvait bénéficier d'une augmentation de la capacité maximale d'OXPHOS mitochondriale spécifique à la masse (adénosine triphosphate (ATP) produite par minute et par gramme de tissu), il devrait alors augmenter l'activité spécifique d'OXPHOS dans le tissu.
Parce que la membrane mitochondriale interne est densément remplie de complexes OXPHOS, il y a peu de potentiel pour augmenter la capacité OXPHOS en augmentant la quantité de protéine OXPHOS dans une quantité donnée de membrane interne. Ainsi, une augmentation de l'activité spécifique d'OXPHOS peut survenir par (1) une augmentation de la quantité de membrane mitochondriale interne contenue dans un volume de mitochondries ou (2) une augmentation du volume de mitochondries dans un volume de tissu. Les deux stratégies peuvent être distinguées par l'analyse morphométrique mitochondriale.
Transport du calcium à travers la membrane mitochondriale interne
La membrane mitochondriale interne (MMI) est moins perméable aux ions et aux petites molécules que la membrane mitochondriale externe (MME). L'activité de la chaîne respiratoire génère un gradient électrochimique à travers la MMI (-180 mV à l'intérieur) qui pilote la synthèse d'ATP et permet l'absorption de Ca2+ par les mitochondries. En conséquence, le traitement avec un découpleur tel que la p-(trifluorométhoxy)-phényl-hydrazone (FCCP), qui effondre le ΔΨm, supprime l'accumulation mitochondriale de Ca2+.
Sur la base de la faible affinité pour le Ca2+ de l'uniporteur mitochondrial, on a initialement supposé que les mitochondries n'affecteraient pas la dynamique du Ca2+ dans des conditions physiologiques, mais agiraient plutôt comme un tampon de faible affinité en cas de surcharge pathologique en Ca2+. Un nouveau rôle des mitochondries dans l'homéostasie du Ca2+ est apparu lorsque le [Ca2+]m a été mesuré directement dans les cellules vivantes au moyen d'une sonde sensible au Ca2+ ciblée sur les mitochondries et basée sur la photoprotéine équorine. Contrairement aux prédictions précédentes, une augmentation rapide de [Ca2+]m a été détectée en réponse à des augmentations physiologiques de [Ca2+]c déclenchées par la libération de Ca2+ du réticulum endoplasmique (RE) induite par un agoniste. Cette divergence apparente a été expliquée par la démonstration que des microdomaines de haut [Ca2+] sont générés dans des sites de proximité étroite entre les mitochondries et le réticulum endoplasmique (RE) et induisent l'ouverture de l'uniporteur.
Myopathie mitochondriale
La myopathie mitochondriale résulte de défauts de la composition du milieu lipidique mitochondrial. La membrane mitochondriale interne (MMI) contient des phospholipides essentiels à la structure et à l'intégrité des protéines de la chaîne respiratoire intégrées. Le syndrome de Barth est un trouble prototypique du métabolisme des phospholipides mitochondriaux avec des anomalies secondaires de la chaîne respiratoire. Ce trouble récessif lié à l'X est causé par des mutations dans TAZ1, codant pour la monolysocardiolipine transacylase, nécessaire au remodelage de la cardiolipine. La maladie se caractérise cliniquement par une myopathie cardiaque et squelettique mitochondriale, une neutropénie cyclique et un retard de croissance avec des anomalies des activités enzymatiques de la chaîne respiratoire.
Un deuxième trouble du milieu lipidique mitochondrial est une myopathie congénitale autosomique récessive avec retard mental et évolution prolongée. La cause est des mutations dans CHKB, codant pour la choline kinase bêta, une enzyme nécessaire à la synthèse de novo de la phosphatidylcholine et de la phosphatidyléthanolamine. Des biopsies musculaires ont révélé des mitochondries étonnamment agrandies qui ont été déplacées vers la périphérie des fibres.
Le syndrome de Sengers, une maladie autosomique récessive qui se manifeste par des cataractes congénitales, une cardiomyopathie hypertrophique, une myopathie squelettique, une intolérance à l'effort et une acidose lactique, est dû à des mutations du gène AGK codant pour l'acylglycérol kinase. Le défaut AGK provoque une carence en acide phosphatidique et une diminution secondaire de l'ANT dans la MMI.
Synonymes, antonymes
Voir tous les synonymes pour "membrane mitochondriale interne".2 synonymes (sens proche) de "membrane mitochondriale interne" :
- membrane interne d'une mitochondrie
- membrane mitochondriale
1 antonyme (sens contraire) :
Les mots ou les expressions apparentés à MEMBRANE MITOCHONDRIALE INTERNE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
L'expression MEMBRANE MITOCHONDRIALE INTERNE est dans la page 2 des mots en M du lexique du dictionnaire.
Mots en M à proximité
membrane cytoplasmique membrane de fécondation membrane intracellulaire membrane lipidique membrane mitochondriale externe membrane mitochondriale internemembrane nictitante membrane nucléaire membrane plasmique membrane séreuse membrane tectoriale
En rapport avec "membrane mitochondriale interne"

Les crêtes mitochondriales (cristae) sont des invaginations internes, aplaties, tubulaires ou discoïdes, qui sont observables à l'intérieur des mitochondries.
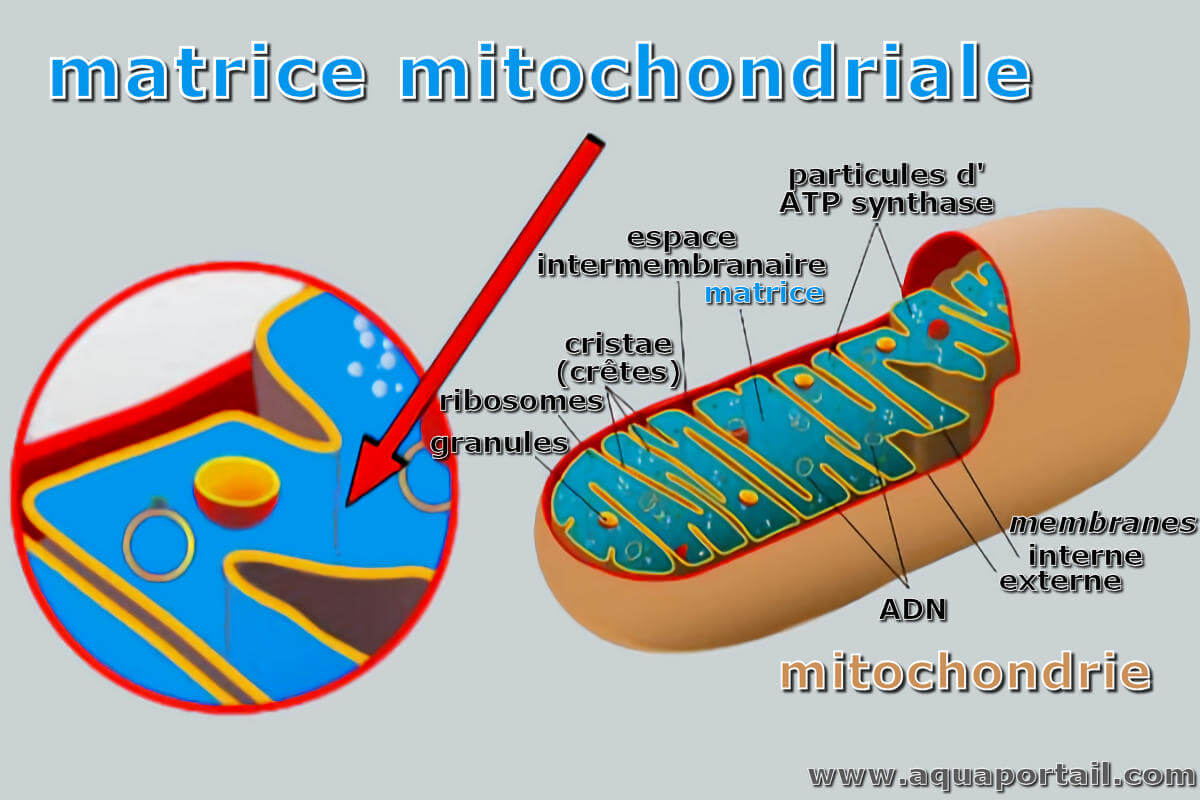
La matrice mitochondriale est l'espace intérieur d'une mitochondrie entouré par la membrane interne mitochondriale, qui contient plusieurs protéines, telles...
membrane mitochondriale externe

La membrane mitochondriale externe entoure complètement la membrane interne d'une mitochondrie, avec un petit espace intermembranaire entre les deux.
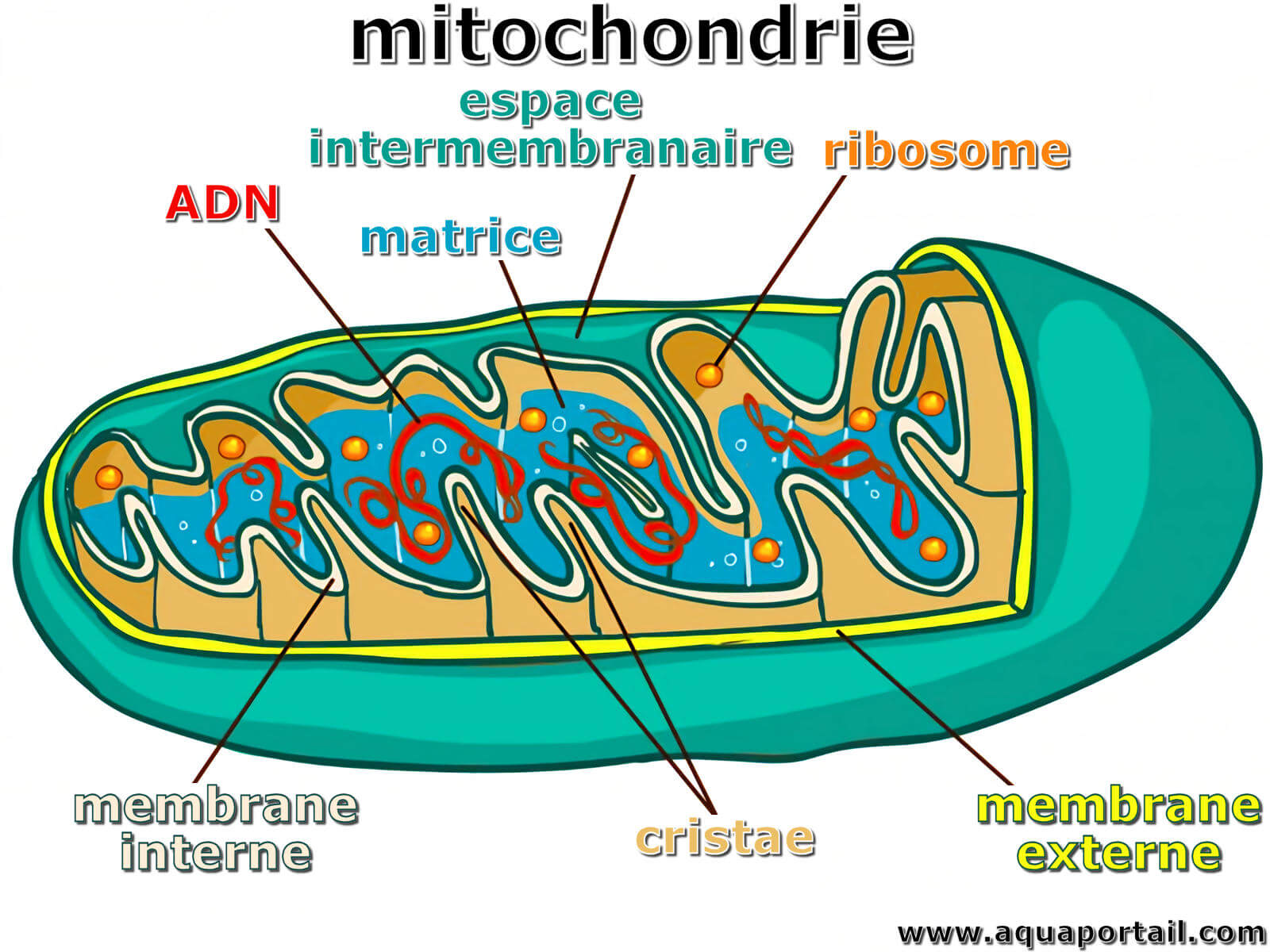
Une mitochondrie est un organite cytoplasmique mobile des cellules eucaryotes qui agit comme une centrale énergétique en produisant de l'ATP...