
Photosystème 1
Définition
Le photosystème 1 (PSI) est un centre réactionnel, un composant essentiel, de la photosynthèse contenant le pigment P700. Il existe un deuxième photosystème : le photosystème 2. Il permet la formation de substances organiques à l'aide de la lumière en tant que source d'énergie présente dans les plantes, les algues, les protistes photosynthétiques et les bactéries photosynthétiques.
Le modèle d'un photosystème 1 (PSI) :
Le photosystème I (premier photosystème, le photosystème 1, FSI), ou plastocyanine-ferrédoxine-oxydoréductase, est le deuxième complexe fonctionnel de la chaîne de transport d'électrons (ETC) des chloroplastes. Il prend un électron de la plastocyanine et, absorbant l'énergie lumineuse, forme un puissant agent réducteur P700, capable de réduire le NADP+ à travers une chaîne de porteurs d'électrons. Ainsi, avec la participation du PSI, une source d'électrons (NADPH) est synthétisée pour des réactions de réduction de carbone ultérieures dans les chloroplastes du cycle de Calvin. De plus, PSI peut effectuer un transport cyclique d'électrons couplé à la synthèse d'ATP, fournissant une synthèse supplémentaire d'ATP dans les chloroplastes.
Explications
Le photosystème 1 est un système de matériaux particulièrement structuré constitué d'un complexe collecteur de lumière et de réaction. Dans le photosystème I, un agent réducteur puissant (NADPH) est formé pour la synthèse de substances organiques à partir de dioxyde de carbone et d'eau, et l'énergie lumineuse est convertie en une source d'énergie appropriée pour cette biosynthèse de l'adénosine triphosphate (ATP).
Propriétés
Le photosystème I (PSI) est un complexe protéique composé de plusieurs protéines membranaires qui catalyse l'oxydation de la plastocyanine et la réduction de la ferrédoxine par des photons absorbés au cours de la photosynthèse. Il se trouve dans les cellules végétales de la membrane thylacoïdienne des chloroplastes et consiste en 15 protéines. Il sert à absorber les photons et à les diriger vers le centre de réaction P 700, complexe moléculaire contenant de la chlorophylle-a et caractérisé par une absorption à une longueur d'onde d'environ 700 nm. Il sert de photosensibilisateur. Après excitation par des photons, un couple de radicaux secondaires se forme en moins de 100 picosecondes, qui se désintègrent à la température ambiante en 300 nanosecondes ou en 300 µs à 77 Kelvin.
Les molécules de chlorophylle a restantes, ainsi que les autres chlorophylles, biliprotéines et caroténoïdes, ont une fonction d'antenne, i. c'est-à-dire qu'ils transfèrent l'énergie rayonnante absorbée vers le centre de réaction. Ils sont disposés de manière à capter la lumière dans la membrane thylacoïdienne en deux couches parallèles (luminal et stromal). Cependant, dans le domaine du transport d'électrons, ils sont disposés dans une pseudo-symétrie C2 orthogonale à la membrane. Le plus important pour la photosynthèse ici est le transfert de l'énergie d'excitation photochimique vers les molécules voisines. Dans ce cas, le spectre de fluorescence de la molécule donneuse doit chevaucher le spectre d'absorption de la molécule réceptrice et les molécules de chlorophylle sont fortement encombrées dans la membrane. Les processus de transfert d'énergie et de transfert d'électrons ont lieu.
À la température ambiante, le spectre de fluorescence de la chlorophylle-a montre une forte bande d'absorption à 685 nm et une plus faible à 740 nm (si les spectres sont déterminés à basse température, des bandes apparaissent à 685, 695 et 720 nm, les deux premiers étant le système pigmentaire II, qui sont affectés au système de pigment I à 720 nm.
La composition en pigment des photosystèmes chez les plantes supérieures est la suivante :
- Photosystème I : carotènes, chlorophylle a, chlorophylle b;
- photosystème II : xanthophylle, chlorophylle a, chlorophylle b.
Les photosystèmes diffèrent non seulement par le spectre d'absorption du pigment dans le centre de réaction, mais également par la composition des pigments d'antenne. Par exemple, le photosystème II est plus riche en chlorophylle b. Le photosystème I est moins sensible aux augmentations de température que le photosystème II.
Composition
Deux protéines du photosystème I, PsaA et PsaB, ont chacune onze hélices transmembranaires et une masse d'environ 80 kDa. Elles forment un hétérodimère et se lient comme cofacteur à environ 80 molécules de chlorophylle-a, à environ 20 β-carotène pour l'absorption de la lumière et à environ six molécules de chlorophylle. Des molécules, deux phylloquinones et un cluster 4Fe-4S servent pour le transfert d'électrons. Les treize autres protéines sont relativement petites, avec des masses comprises entre quatre et dix-huit kilodaltons.
Transport de photons et d'électrons
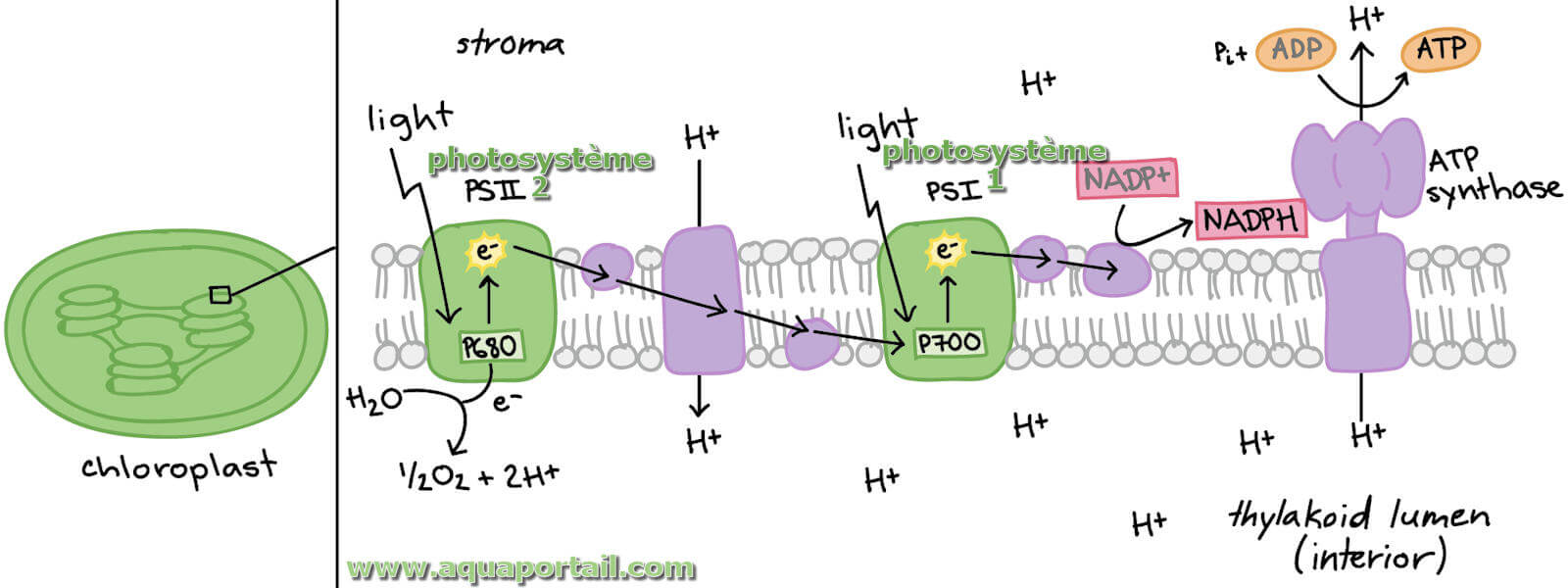
Les substances des photosystèmes agissant comme antennes sont regroupées dans un complexe collecteur de lumière (LHC). Grâce à cela, la surface absorbant la lumière est augmentée. Les centres de réaction sont les systèmes rédox actuels du transport d'électrons photochimique. Le centre de réaction P700 du photosystème I est un agent réducteur puissant (le potentiel de réduction standard à l'état fondamental est d'environ +0,45 V; à l'état excité photochimiquement, la valeur est de -0,6 V). Il absorbe deux photons et agit ensuite comme donneur d'électrons primaire, en transférant un total de deux électrons sur le récepteur primaire A, une protéine fer-soufre, avec deux excitations photoniques. Le récepteur primaire est capable de réduire la ferrédoxine liée à la membrane. Cela signifie que la ferrédoxine est ici l'accepteur d'électrons. De là, les électrons sont transférés à une enzyme contenant le FAD.
Cela catalyse la réaction suivante : NADP+ effectue la transition vers NADPH + H+ en prenant deux protons et deux électrons.
Le produit de réaction du photosystème I est l'ATP de stockage d'énergie : dans une réaction de phosphorylation, l'ATP est formé d'ADP et de phosphate.
L'ATP est le fournisseur d'énergie pour les processus intervenant dans l'obscurité (respiration). La compensation d'électrons a lieu dans le cours ultérieur en coopération avec le photosystème II en jouant le rôle de plastocyanine donneuse. C'est une protéine aux propriétés rédox qui contient deux ions de cuivre liés dans son centre actif. Celui-ci reçoit ses électrons du complexe cytochrome b, qui est à son tour réduit par la plastoquinone. La plastoquinone elle-même se transforme en plastoquinol lors de l'absorption d'électrons. Ceci ferme la chaîne et équilibre le bilan des électrons. Dans la réaction globale de la photosynthèse, de l'oxygène est également produit, notamment dans la réaction à la lumière dans le photosystème II, formé à partir de l'eau. Le carotène présent dans le photosystème I en même temps que la chlorophylle absorbe et élimine l'oxygène singulet formé, qui est toxique pour les feuilles de la plante.
Différentes réactions dans l'obscurité peuvent suivre, par exemple, l'utilisation enzymatique du carbone lié, qui transforme la photosynthèse en un processus cyclique appelé cycle de Calvin, son principal découvreur. Globalement, le processus intervenant dans le photosystème I peut être classé dans une chaîne de transport d'électrons. On peut noter que la photosynthèse est une photoréaction avec un rendement quantique supérieur à 90 %.
Le gradient de proton nécessaire à la production d'ATP peut être généré non seulement via la chaîne de transport d'électrons linéaire, mais également via un transport d'électrons cyclique dans le photosystème I.
Différences par rapport au photosystème 2 (PSII)
La fonction principale du photosystème II est la génération d'un agent oxydant puissant, qui initie l'oxydation de l'eau et le transfert de ses électrons sur le support membranaire. La fonction principale du photosystème I est de remplir d'énergie ces électrons de bas niveau afin de les utiliser pour restaurer le NADP+. Puisque l'énergie du processus total est trop élevée pour être effectuée dans le même centre de réaction, il est apparu au cours de l'évolution deux photosystèmes qui effectuent séparément différentes parties de cette réaction. Leurs fonctions spécifiques et déterminent les caractéristiques de leur structure.
Le photosystème I est symétrique, c'est-à-dire qu'il a deux branches de transport électronique, ce qui le rend beaucoup plus rapide, tandis que le photosystème II est asymétrique et n'a qu'une branche active, ce qui ralentit le transport des électrons, mais le rend plus maniable. Les deux photosystèmes diffèrent de manière significative dans la structure des antennes, des sous-unités supplémentaires, des méthodes de régulation et leur position dans la membrane.
Ainsi, le photosystème I possède une antenne intégrale, dont les chlorophylles sont situées directement sur les protéines principales du complexe, A et B, tandis que dans le photosystème II, elles sont placées sur les protéines externes CP47 et CP43. En termes de nombre de petites sous-unités régulatrices supplémentaires de PSII, il dépasse de loin PSI, ce qui est associé à la nécessité de réguler avec précision le processus d'oxydation de l'eau, qui est potentiellement extrêmement dangereux pour la cellule. Cela explique également la distribution inhomogène des photosystèmes dans la membrane thylacoïdienne : alors que PS I est principalement située dans les membranes marginale, terminale et stromale, PSII se trouve presque complètement dans la région des membranes appariées, ce qui offre à la cellule une protection supplémentaire contre les formes d'oxygène actif produites par celle-ci.
La principale différence entre le photosystème II et le photosystème I est la présence d'un grand domaine faisant face à la lumière, qui contient un cluster de manganèse et les protéines protectrices qui l'entourent. C'est ici que commence le processus d'oxydation photochimique de l'eau, accompagné de la libération d'oxygène et de protons.
Histoire de la découverte
Le transport d'électrons non cyclique commence par le fait que la grappe de manganèse du photosystème II oxyde l'eau pure en restaurant le pool de plastoquinones. En outre, le complexe cytochrome b6f (plastoquinolplasocyanine réductase) oxyde les plastoquinones et un électron par plastocyanine est transféré au photosystème I où il est utilisé pour la synthèse de NADPH. La violation de la logique formelle dans les noms de photosystème est due au fait que le photosystème I a été découvert avant le photosystème II.
Les premières données indiquant l'existence de FSI sont apparues dans les années 1950, mais à cette époque, personne ne pouvait encore comprendre l'importance de ces découvertes. L'idée de l'existence de deux photosystèmes dans les chloroplastes est apparue dans les années 1940 déjà sur la base des expériences du laboratoire de R. Emerson, qui avait découvert l'effet d'une diminution du rendement quantique de la photosynthèse lorsque les chloroplastes étaient éclairés avec une lumière rouge monochromatique (λ > 680 nm), et une augmentation du quantum. sortie lors de l'ajout d'une lumière d'une longueur d'environ 650 nm, ce qui excitait le PSII (l'effet appelé Emerson). Il convient également de mentionner le signal de RPE induit par la lumière découvert par Comonner en 1956, appelé signal I. Par hasard, les signaux I et II provenaient respectivement des interfaces PSI et PSII. Ce n'est qu'en 1960 que Louis Duisens a proposé le concept de photosystème I et de photosystème II, et la même année, Fei Bendall et Robert Hill ont organisé les résultats des découvertes précédentes dans une théorie cohérente des réactions séquentielles de la photosynthèse. L'hypothèse de Hill et Bendall a ensuite été confirmée par les expériences de Duysens et de Witt en 1961.
Après cela, des tentatives systématiques ont été faites pour isoler physiquement le photosystème I, afin de déterminer sa structure tridimensionnelle et sa structure fine. En 1966, les recherches dans ce domaine ont commencé à se développer : Anderson et Boardman ont soumis les membranes de chloroplaste à des ultrasons suivis par un traitement à la digitonine, Vernon a utilisé le triton X-100 et Ogawa a utilisé du dodécyl sulfate. Cependant, les premiers extraits obtenus contenaient des impuretés de complexes collecteurs de lumière, ainsi que des cytochromes f et b6. Il a fallu beaucoup de temps pour comprendre que les extraits étaient un mélange.
En 1968, Reed et Clayton ont été capables d'isoler le centre réactionnel du photosystème I de bactéries pourpres, ce qui a grandement incité à l'étude de la photosynthèse oxygénique. Cependant, la question restait ouverte : quel était le véritable centre réactionnaire choisi, quels étaient les complexes d'antennes et quelles étaient les sous-unités supplémentaires. Pendant longtemps, la sélection effective du centre de réaction du photosystème I est restée un problème non résolu. Au final, il s'est avéré qu'il était plus facile de le faire pour les cyanobactéries, car elles manquaient d'antennes externes intégrées à la membrane. Après de nombreuses tentatives avec différentes espèces, il s'est avéré que les espèces les plus prometteuses à cet égard sont des représentants de Synechocystis et de Synechococcus, car le photosystème I, isolé de Thermosynechococcus elongatus, constituait un centre de réaction très stable, adapté à la cristallisation et à l'analyse par rayons X.
Synonymes, antonymes
Voir tous les synonymes pour "photosystème 1".1 synonyme (sens proche) de "photosystème 1" :
1 antonyme (sens contraire) :
Les mots ou les expressions apparentés à PHOTOSYSTÈME 1 sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
Le mot PHOTOSYSTEME 1 est dans la page 4 des mots en P du lexique du dictionnaire.
Mots en P à proximité
photosynthèse en C4 photosynthèse oxygénique photosynthétat photosynthétique photosystème photosystème 1photosystème 2 phototactique phototactisme phototaxie phototransduction
En rapport avec "photosystème 1"
centres réactionnels de la photosynthèse

Les centres réactionnels de la photosynthèse sont des centres privilégiés de la photosynthèse où des réactions d'oxydoréduction interviennent.
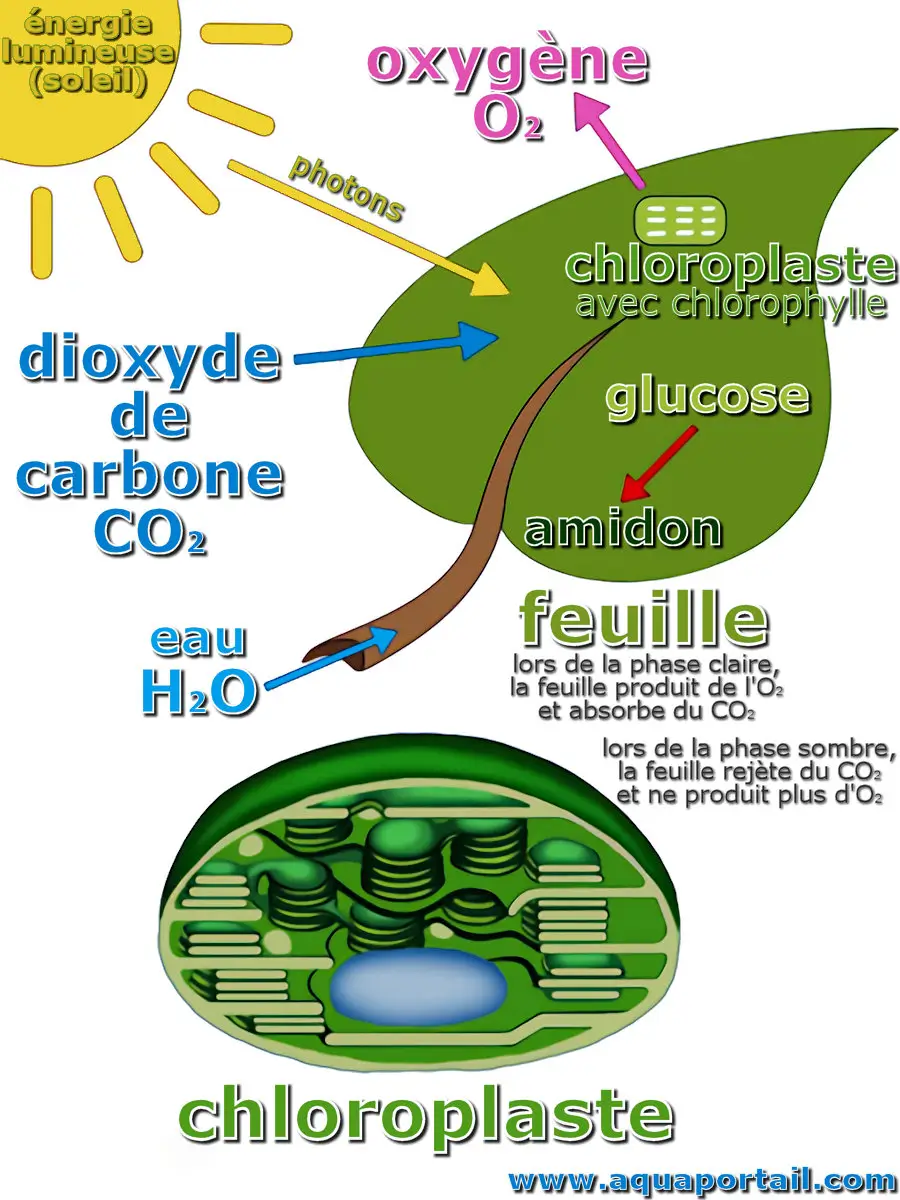
La photosynthèse est le bioprocédé de conversion de substances organiques (CO₂, eau) en sucre tout en dégageant de l'oxygène (O₂) grâce à l'énergie photonique...
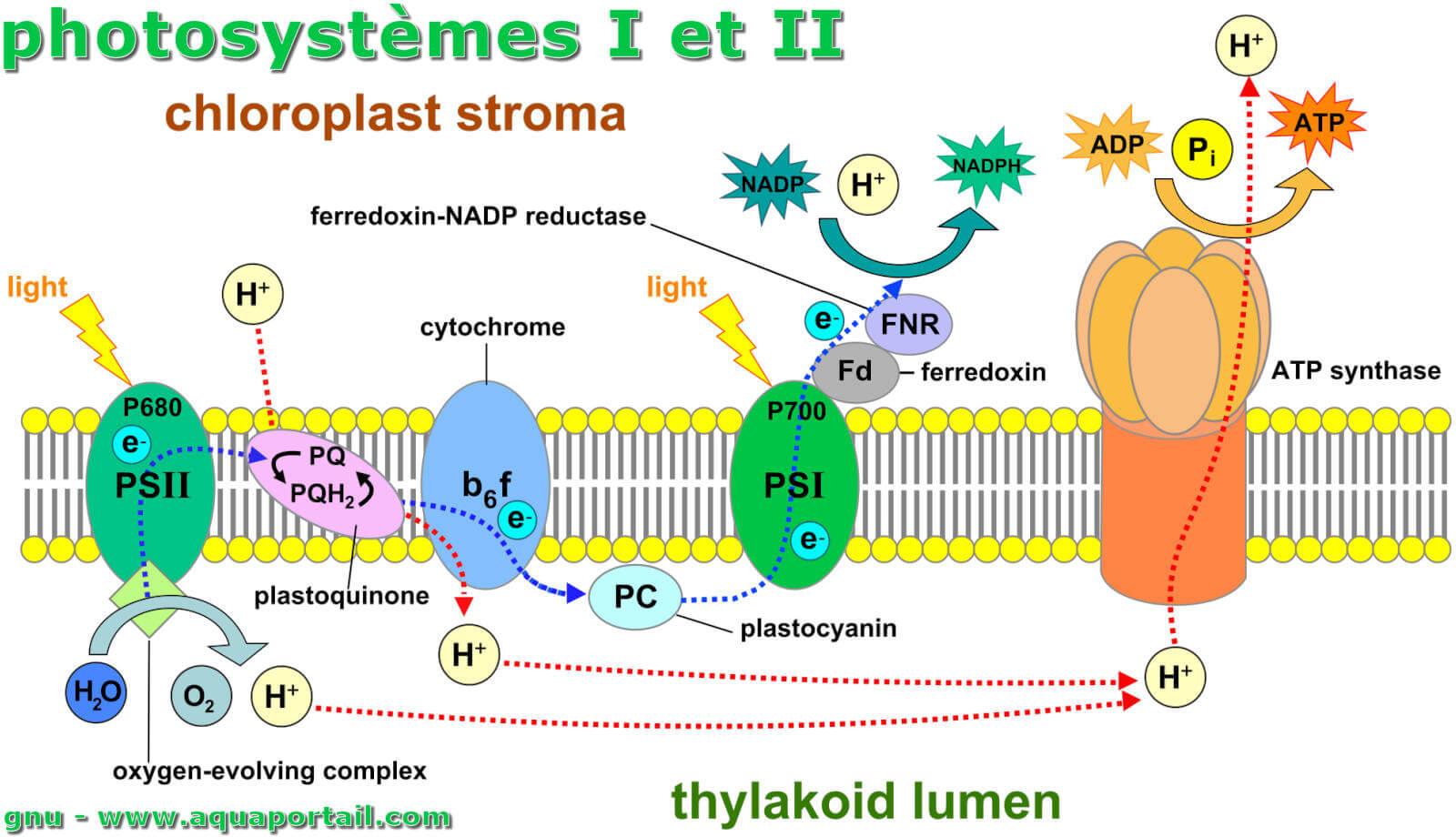
Un photosystème qualifie le centre réactionnel de la photosynthèse, chacun des complexes protéiques des cellules photosynthétiques formés d'une antenne...
Le photosystème II (PSII) est un supercomplexe de protéines membranaires qui exécute la réaction initiale de photosynthèse chez les plantes.