
Brassage génétique
Définition
Le brassage génétique correspond aux recombinaisons génétiques au sein d'une population entière et sur plusieurs générations. Une recombinaison génétique consiste en tout réarrangement physique de la succession des loci. La recombinaison génétique permet le brassage génétique dans un flux.
Modèle de brassage génétique avec recombinaison méiotique :
Un modèle actuel de brassage génétique avec recombinaison méiotique, initié par une rupture ou un gap à double brin, suivi par l'appariement avec un chromosome homologue et l'invasion de la chaîne pour initier le processus de réparation recombinatoire. La réparation de la séparation peut conduire à un croisement ou non des régions. On pense que la recombinaison de croisement (crossover) intervient à travers le modèle "Double Holliday Junction (DHJ)", illustré sur la droite, ci-dessus. On pense que les recombinants sans croisement sont produits principalement par le modèle de recuit de brin dépendant de la synthèse (SDSA), illustré ci-dessus à gauche. La plupart des événements de recombinaison semblent être du type SDSA.
Explications
La recombinaison en biologie est le réarrangement du matériel génétique (ADN, ARN) dans les cellules et, dans un sens plus étroit, l'échange d'allèles. La recombinaison conduit à de nouvelles combinaisons de gènes et de caractéristiques. La recombinaison et la mutation entraînent une variabilité génétique au sein d'une population. À son tour, la variabilité génétique est la base de l'adaptation aux changements des conditions environnementales dans le processus de microévolution.
Voir aussi les transposons (gènes sauteurs) et la transposase.
Fonctionnement de la recombinaison génétique
La recombinaison génétique ou le brassage est le processus par lequel un brin de matériel génétique (généralement de l'ADN, mais qui peut aussi être de l'ARN) est coupé puis attaché à une molécule différente de matériel génétique. Chez les eucaryotes, la recombinaison survient fréquemment pendant la méiose de la reproduction sexuée (le processus par lequel les eucaryotes génèrent des gamètes), comme une réticulation chromosomique entre des chromosomes appariés. Ce processus conduit à la progéniture ayant des combinaisons de gènes différents de ceux de leurs parents et peut produire des allèles chimères.
En biologie de l'évolution, on pense que ce mélange de gènes a plusieurs avantages, notamment qu'il permet aux organismes de se reproduire sexuellement et d'éviter le cliquet de Muller. Chez les vertébrés, il existe également un type spécial de recombinaison génétique dans les cellules B et T du système immunitaire, appelé recombinaison V(D)J, qui est responsable de la génération de la variabilité énorme des anticorps et des récepteurs des lymphocytes T nécessaires à la réponse immunitaire La recombinaison site-spécifique est un type particulier de recombinaison homologue qui intervient dans des régions spécifiques, courtes et homologues, existant dans les deux fragments à recombiner. Il est généralement typique des virus, qui l'utilisent pour entrer dans le génome de l'hôte.
En biologie moléculaire, la "recombinaison" fait également référence à la recombinaison artificielle et délibérée de différents fragments d'ADN, souvent issus d'organismes différents, créant ce qu'on appelle l'ADN recombinant (ADNr).
Il existe plusieurs types de recombinaison génétique dans les cellules eucaryotes :
- La recombinaison homologue (également appelée recombinaison générale) survient au cours de la prophase I de la méiose et se déroule entre de longues régions d'ADN dont les séquences sont homologues, c'est-à-dire hautement similaires mais non identiques.
- La recombinaison peut intervenir entre des séquences d'ADN qui ne contiennent pas de séquences homologues. Ceci est connu comme une recombinaison non homologue. Elle survient rarement chez les procaryotes et les levures, mais il est plus commun dans les cellules de mammifères.
- La recombinaison spécifique au site intervient en rompant et en rejoignant ensuite des régions d'homologie courte et spécifique de deux ADN différents, ou au sein d'une même molécule. Il existe dans les virus (par exemple, le bactériophage T4) et dans les plasmides.
Mécanisme du brassage génétique
Le brassage génétique est catalysé par plusieurs enzymes. Les recombinases sont les enzymes clés qui catalysent l'étape de transfert du brin pendant la recombinaison. La RecA (recombinase trouvée dans Escherichia coli) est responsable de la réparation des cassures double brin de l'ADN. Chez les levures et autres organismes eucaryotes, il est nécessaire d'utiliser deux recombinases pour la réparation des cassures double brin dans l'ADN. La protéine RAD51 est nécessaire pour la recombinaison mitotique et méiotique, tandis que la protéine de réparation de l'ADN, DMC1, est spécifique à la recombinaison méiotique. Dans les archées, l'orthologue de la protéine bactérienne RecA est RadA.
Génie génétique
En génie génétique, la recombinaison peut également se référer à la recombinaison artificielle et délibérée de fragments d'ADN, souvent issus de différents organismes, créant ce qu'on appelle l'ADN recombinant. Un exemple de recombinaison génétique est le gène de ciblage, qui peut être utilisé pour ajouter, supprimer ou modifier les gènes d'un organisme. Cette technique est importante pour les chercheurs biomédicaux puisqu'elle leur permet d'étudier les effets de gènes spécifiques. Des techniques basées sur la recombinaison génétique sont également appliquées dans l'ingénierie des protéines pour développer de nouvelles protéines d'intérêt biologique.
Recombinaison méiotique
Les modèles moléculaires de la recombinaison méiotique ont évolué au fil des ans, fournissant ainsi une grande quantité de preuves. La compréhension fondamentale du mécanisme de la recombinaison méiotique est cruciale pour résoudre le problème de la fonction adaptative du sexe, une question qui n'a pas été résolue en biologie. Un modèle qui reflète la compréhension actuelle a été présenté par Anderson et Sekelsky 12 et est décrit dans la première figure de cet article. La figure montre que deux des quatre chromatides présentes dans la méiose (prophase I) sont appariées et peuvent interagir. La recombinaison, dans cette version du modèle, est initiée par une cassure à double brin qui est montrée dans la molécule d'ADN (chromatide). Cependant, d'autres types de dommages à l'ADN peuvent également déclencher une recombinaison. Par exemple, la réticulation inter-brins (causée par l'exposition à un agent de réticulation tel que la mitomycine C) peut être réparée par HRR.
Comme indiqué dans la première figure ci-dessus, deux types de produits recombinants sont produits. Sur le côté droit il y a un type " cross over " (CO), où les chromatides des chromosomes homologues sont échangés et sur le côté gauche, un type "no crossover" (NCO) où les chromatides ne sont pas échangées. Le type de recombinaison CO implique la formation intermédiaire de deux " Holliday Jonctions " indiquées dans la partie inférieure droite de la figure par deux structures en forme de X dans chacune desquelles il y a un échange de brins individuels entre les deux chromatides participantes. Cette route est étiquetée dans la figure sous le nom de "Double Holliday Junction (DHJ)".
Les recombinants NCO (illustrés à gauche sur la figure) sont produits par un procédé appelé recuit de brin dépendant de la synthèse (SDSA). Les événements de recombinaison du type NCO / SDSA semblent être plus communs que le type CO/DHJ. La voie NCO/SDSA contribue peu à la variation génétique puisque les bras des chromosomes flanquant l'événement de recombinaison restent dans la configuration parentale. Par conséquent, les explications de la fonction adaptative de la méiose qui se concentrent exclusivement sur le croisement sont insuffisantes pour expliquer la plupart des événements de recombinaison.
Réparation recombinante
La réparation recombinante nécessite la présence d'une séquence identique ou presque identique qui est utilisée comme matrice pour réparer la rupture. La machinerie enzymatique responsable de ce processus est presque identique à la machinerie responsable du croisement des chromosomes pendant la méiose. Cette voie permet de réparer un chromosome endommagé en utilisant une chromatide soeur (disponible en G2 après la réplication de l'ADN) ou un chromosome homologue en tant que matrice.
Au cours de la mitose et de la méiose, les lésions de l'ADN provoquées par une variété d'agents exogènes (par exemple la lumière UV, les rayons X, les agents de réticulation chimiques) peuvent être réparées par réparation recombinante homologue (HRR). Ces résultats suggèrent que les dommages à l'ADN qui résultent de processus naturels, tels que l'exposition à des espèces réactives de l'oxygène (ROS) qui sont des sous-produits du métabolisme normal, sont également réparés par HRR. Chez les humains et les rongeurs, les problèmes dans les gènes nécessaires pour la HRR pendant la méiose provoquent l'infertilité. Tandis que dans le problème humain dans les gènes tels que BRCA1 et BRCA2, ils augmentent le risque de cancer.
Chez les bactéries, la transformation est un processus de transfert de gènes qui se déroule normalement entre des cellules individuelles de la même espèce bactérienne. La transformation implique l'intégration de l'ADN du donneur dans le chromosome receveur par recombinaison. Ce processus semble être une adaptation pour la réparation des dommages à l'ADN du chromosome récepteur par HRR. La transformation peut apporter un bénéfice aux bactéries pathogènes en permettant la réparation des dommages à l'ADN.
Lorsque deux ou plusieurs virus, chacun avec des dommages génomiques létaux, infectent la même cellule hôte, les génomes du virus peuvent souvent être appariés entre eux et sont réparés par HRR afin de produire une descendance viable. Ce processus appelé réactivation de la multiplicité a été étudié dans les bactériophages T4 et lambda 14 ainsi que dans plusieurs virus pathogènes. Dans le cas des virus pathogènes, la réactivation de la multiplicité peut être un avantage adaptatif pour le virus, car elle permet la réparation des dommages à l'ADN causés par l'exposition au milieu oxydant produit pendant l'infection de l'hôte.
Synonymes, antonymes
Voir tous les synonymes pour "brassage génétique".2 synonymes (sens proche) de "brassage génétique" :
- recombinaison génétique
- recombinaison méiotique
0 antonyme (sens contraire).
Les mots ou les expressions apparentés à BRASSAGE GÉNÉTIQUE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
L'expression BRASSAGE GENETIQUE est dans la page 4 des mots en B du lexique du dictionnaire.
Mots en B à proximité
brannérite bras bras de mer bras deltaïque bras-mort brassage génétiqueBrassicacées Brassicales brassinolide brassinostéroïde bréchet
En rapport avec "brassage génétique"
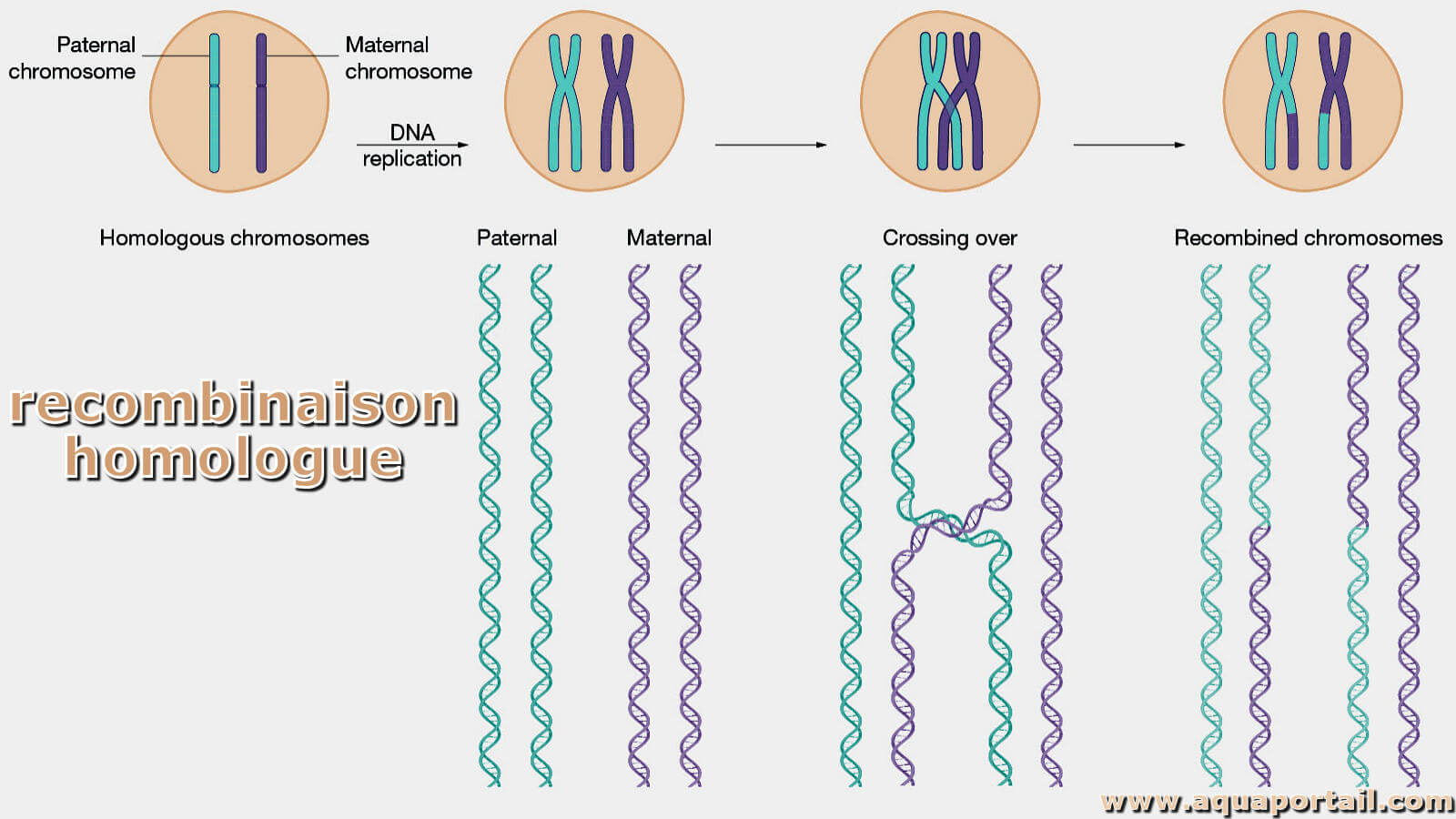
La recombinaison est une nouvelle combinaison, inverse de la dissociation. En génétique, elle s'effectue par conjugaison, transduction ou transformation.
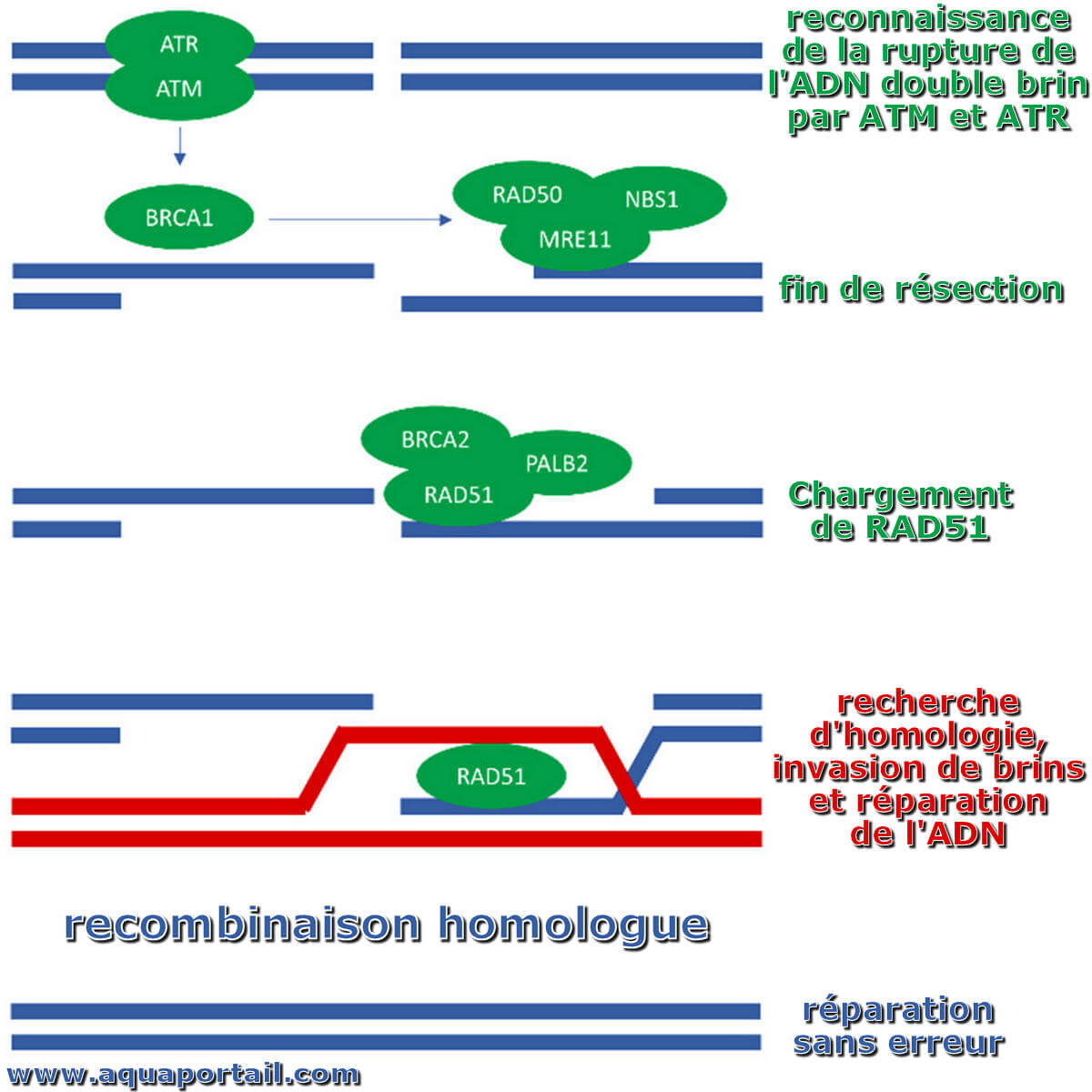
La recombinaison homologue est une voie de réparation de la rupture double brin de l'ADN qui protège le génome de l'instabilité chromosomique.

Une action génétique additive identifie un type d'action génétique où chacun des deux allèles contribue de la même façon à la production de phénotypes...

Une action génétique en dominance est un type d'action génétique où un allèle l'emporte sur l'autre, parfois de façon complète mais d'autres fois, de façon...