
Motoneurone
Définition
Un motoneurone (MN, neurone moteur) est une cellule énergivore sans lequel la vie complexe ne serait pas possible. Toute interférence avec la production d'énergie mitochondriale des motoneurones est généralement préjudiciable à sa survie.
Trois types de neurones (avec motoneurone) :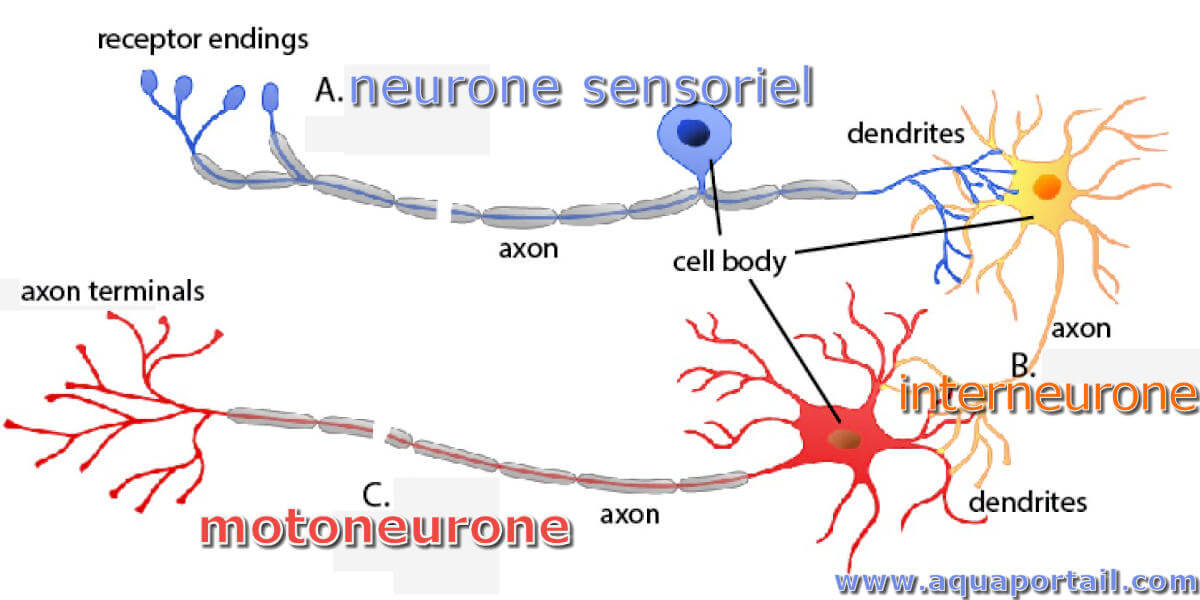
Trois types de neurones et leurs relations spatiales. La recherche sur les motoneurones pourrait élargir la compréhension des cellules nerveuses.
Explications
Les motoneurones forment un groupe diversifié de cellules sans lesquelles la vie complexe ne serait pas possible. Les motoneurones sont responsables de l'intégration des signaux du cerveau et des systèmes sensoriels pour contrôler les mouvements volontaires et involontaires.
Bien que les motoneurones puissent être divisés en sous-ensembles crâniens et rachidiens, les motoneurones rachidiens sont les plus importants des neurones moteurs car ils sont une cible clé des maladies et des blessures. En tant que tels, les motoneurones sont au centre des efforts de régénération visant à alléger ces fardeaux de santé publique.
Au cours de la gastrulation et de la neurulation tardives, la moelle épinière en développement, appelée tube neural, est structurée en domaines progéniteurs distincts. Les motoneurones sont spécifiés à partir de progéniteurs dans le tube neural ventral. Une fois spécifiés, les motoneurones nouvellement nés sont ensuite spécifiés en colonnes, pools et sous-types, formant une topographie unique. À partir de ces colonnes et pools, les axones atteignent leurs cibles sous divers signaux de guidage.
Tous les motoneurones sont des cellules cholinergiques qui s'intègrent au circuit de commande du moteur, au système sensoriel et à leurs cibles périphériques pour contrôler le mouvement. Les MN sont uniques en ce sens que leurs cibles se situent en dehors du système nerveux central (SNC), ce qui signifie qu'ils nécessitent de nouvelles méthodes pour les rechercher et synapser dessus. La différenciation et du développement de motoneurones sont des événements de signalisation, les marqueurs des facteurs de transcription et la matrice extracellulaire (E CM) en ce qui concerne le développement de MN.
Ces cellules sont la cible de maladies permanentes et souvent mortelles, notamment la sclérose latérale amyotrophique, l'amyotrophie spinale, la sclérose en plaques et des lésions telles que les lésions de la moelle épinière. Ce n'est qu'en comprenant comment ces cellules progressent dans le développement que nous pouvons comprendre comment traiter ces maladies qui ont actuellement peu d'espoir de guérison.
De plus, en décodant les événements et acteurs majeurs du développement, on peut mieux les récapituler in vitro pour la thérapie de remplacement cellulaire, ou capter les principes sous-jacents à la régénération chez l'adulte. Compte tenu de l'importance croissante de l'interaction MN-glie dans un certain nombre de maladies neurodégénératives, nous discuterons également en détail de la spécification initiale des cellules précurseurs d'oligodendrocytes (OPC), car elles partagent un progéniteur commun avec les motoneurones.
Caractéristiques
Le motoneurone a des dendrites, un corps cellulaire et un axone. Les motoneurones sont de grosses cellules situées dans la corne ventrale de la moelle épinière. Ils ont un certain nombre de processus appelés dendrites qui transmettent des signaux au motoneurone. Le motoneurone possède également un grand processus, l'axone, qui relie le motoneurone à une extrémité à une fibre musculaire à l'autre. Les potentiels d'action se déplacent le long de l'axone de sorte que l'activité dans le motoneurone modifie l'activité dans le muscle.
Les axones des neurones peuvent être myélinisés ou non myélinisés. La myéline fait référence à une gaine qui recouvre l'axone, mais pas entièrement. Dans le système nerveux périphérique, les cellules de Schwann fabriquent la myéline en s'enroulant autour de l'axone, formant une structure multicouche de plusieurs membranes cellulaires de la cellule de Schwann. Dans le système nerveux central, les cellules oligodendrogliales fabriquent la myéline. La gaine n'est continue ni dans le système nerveux périphérique ni dans le système nerveux central. À la fin de chaque cellule de Schwann, il y a un vide dans la myéline, comme un noeud. Cet écart s'appelle le noeud de Ranvier.
Comme toutes les cellules, le motoneurone possède un noyau situé dans son corps cellulaire ou soma. Le soma est aussi parfois appelé le péricaryon de la racine grecque "peri" signifiant "autour" ou "entourant" et "karyon" signifiant "noix" ou "noyau", et se référant au noyau. Il n'y a qu'un seul noyau dans le motoneurone et c'est le site de la transcription de l'ARNm.
Motoneurone de survie
Le motoneurone de survie (SMN) forme un complexe avec de nombreuses autres protéines appelées gémines.59 Le complexe SMN, composé de gémines 2 à 8 et non déchirées, se trouve dans le cytoplasme.59–66 Le complexe SMN fonctionne en plaçant l'anneau protéique Sm sur de petits ARN nucléaires (ARNsn, petit ARN interférent) pour former une petite protéine ribonucléaire nucléaire (PRNsn) qui, avec le complexe SMN, est transportée dans le noyau. Les snRNP sont essentiels pour l'épissage correct de tous les gènes.63,65 Ainsi, la perte complète de SMN entraîne la létalité, car les PRNsn sont essentielles à la survie de toute cellule. Outre la fonction de SMN dans l'assemblage de ARNsn, SMN fonctionne également dans l'assemblage des complexes Lsm10, Lsm11 et Sm sur U7 ARNsn. Le U7 PRNsn traite l'extrémité 3′ du message d'histone. Ces deux fonctions ont été clairement démontrées et ont des dosages biochimiques qui peuvent être utilisés pour déterminer l'activité dans divers échantillons. En plus de cette fonction d'assemblage, il a été proposé qu'un motoneurone de survie fonctionne potentiellement dans l'assemblage d'autres complexes de protéines ribonucléaires nucléaires.
Signaux entrants
Un corps cellulaire de motoneurone est situé dans la moelle épinière et sa fibre (axone) fait saillie à l'extérieur de la moelle épinière pour contrôler directement ou indirectement les organes effecteurs, principalement les muscles et les glandes. Il existe des motoneurones supérieurs et des motoneurones inférieurs, le type de cellule décrit précédemment étant un motoneurone inférieur. Les axones des motoneurones inférieurs sont des fibres nerveuses efférentes qui transmettent les signaux de la moelle épinière aux effecteurs. Les motoneurones supérieurs sont des interneurones corticospinaux qui proviennent du cortex moteur du cerveau et descendent dans la moelle épinière où ils activent les motoneurones inférieurs via les synapses. Le terme "motoneurone (neurone moteur)" est généralement limité aux nerfs efférents qui innervent les muscles, à savoir les motoneurones inférieurs.
Le glutamate est le principal neurotransmetteur excitateur du système nerveux central (SNC). Dans des conditions physiologiques normales, le glutamate est libéré des motoneurones présynaptiques via une exocytose dépendante de Ca2+ pour activer les récepteurs exprimés sur les motoneurones post-synaptiques.
Les motoneurones inférieurs (postsynaptiques) peuvent mourir en raison d'une stimulation excessive causée par la liaison du glutamate à leurs récepteurs de glutamate. Les mécanismes moléculaires et déclencheurs de ce type de lésion neuronale (excitotoxicité du glutamate) dans la SLA ne sont pas encore bien établis. La perte de fonction du transporteur glial de recapture du glutamate (SLC1A2, GLT1) ou de l'enzyme DAO peut déclencher une excitotoxicité du glutamate. Dans les modèles SLA, l'expression de SLC1A2 est faible et entraîne l'inhibition de la recapture efficace du glutamate par les astrocytes
Naissance
Les motoneurones sont parmi les premières cellules nerveuses à naître, c'est-à-dire à terminer leur dernier tour de synthèse d'ADN. Peu de temps après la naissance des cellules, les motoneurones commencent à étendre un axone qui quitte la moelle épinière dans les racines ventrales naissantes. Chez les mammifères, les motoneurones qui innervent un muscle individuel sont généralement regroupés dans une colonne orientée longitudinalement qui s'étend sur deux à trois segments de la colonne vertébrale.
Croissance des axones et identité des motoneurones
Les axones moteurs immatures quittent la moelle épinière avant même la formation de leurs muscles cibles. Au cours de leur croissance, les axones sélectionnent des chemins qui mènent aux muscles qu'ils sont destinés à innerver. La capacité d'un motoneurone immature à prendre de telles décisions indique qu'il a une certaine connaissance de son identité, et que différents motoneurones ont donc des identités différentes. Chez l'adulte, les motoneurones qui innervent les fibres musculaires non fatigables à contraction lente ont tendance à être riches en enzymes oxydatives qui peuvent être visualisées par des techniques histochimiques appropriées. Bien avant la formation des premiers NMJ, les motoneurones embryonnaires diffèrent déjà par leurs profils enzymatiques oxydatifs. Cela soutient l'idée que les motoneurones qui innervent un seul muscle diffèrent dans leurs propriétés et que ces différences surviennent avant toute interaction avec le muscle.
Une fois que le contact avec la masse prémusculaire appropriée a été établi, mais pas avant, les axones se ramifient largement. Chez les rats et les souris, les contacts fonctionnels avec les muscles des membres nouvellement formés sont d'abord présents autour du jour embryonnaire 14 (E14), une semaine avant la naissance. Un stade similaire existe chez l'homme vers la neuvième semaine de gestation.
Libération d'acétylcholine des cônes de croissance
Les terminaux des motoneurones cultivés peuvent libérer de l'acétylcholine (ACh), le transmetteur chimique des jonctions neuromusculaires, avant même qu'ils n'entrent en contact avec le muscle. Cela suggère que les motoneurones in vivo synthétisent également l'ACh et possèdent les spécialisations nécessaires pour sa libération dépendante de l'activité à un stade précoce de leur développement.
Cytologie du système nerveux
Les motoneurones forment la division efférente du SNP. Il y en a environ 500 000, transportant des informations du SNC aux effecteurs périphériques dans les tissus périphériques et les systèmes d'organes. Les fibres efférentes sont les axones des motoneurones et transportent les données du SNC. Les deux principaux systèmes efférents sont le système nerveux somatique (SNS) et le système nerveux autonome viscéral (SNA). Le système nerveux somatique comprend les motoneurones somatiques, qui innervent les muscles squelettiques. Le SNS est sous contrôle conscient. Les corps cellulaires des motoneurones somatiques se trouvent dans le SNC. Les axones voyagent à travers les nerfs périphériques, innervant les fibres musculaires squelettiques aux jonctions neuromusculaires.
Le SNA n'est pas sous contrôle conscient, les motoneurones viscéraux stimulant tous les effecteurs périphériques à l'exception des muscles squelettiques. Ils innervent les muscles cardiaques et lisses, le tissu adipeux et les glandes. Les axones moteurs viscéraux du SNC innervent d'autres motoneurones viscéraux dans les ganglions autonomes périphériques. Les corps cellulaires des neurones innervent et contrôlent les effecteurs périphériques. Les fibres préganglionnaires sont des axones qui s'étendent du SNC aux ganglions autonomes. Les fibres postganglionnaires sont des axones qui relient les cellules ganglionnaires aux effecteurs périphériques.
Développement de la moelle épinière
Le développement des motoneurones résulte de la polarisation ventrale du tube neural. Les motoneurones et les cellules ganglionnaires de la racine dorsale apparaissent autour de E27. Les cellules exprimant Nkx6.1, Olig2 et Pax6 forment le sous-domaine dit des motoneurones. Les cellules de la crête neurale donnent naissance aux neurones sensoriels du système nerveux périphérique, aux neurones postganglionnaires, aux cellules de Schwann, aux cellules satellites des ganglions de la racine dorsale et des ganglions autonomes, et aux cellules endocrines des paraganglions, y compris les cellules adrénergiques des surrénales. Les molécules impliquées dans l'induction des cellules de la crête neurale comprennent les protéines morphogénétiques osseuses et Wnt6
Synonymes, antonymes
1 synonyme (sens proche) de "motoneurone" :
0 antonyme (sens contraire).
Les mots ou les expressions apparentés à MOTONEURONE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
Le mot MOTONEURONE est dans la page 5 des mots en M du lexique du dictionnaire.
Mots en M à proximité
morula moteur moléculaire motif CpG motiline motilitémotoneuronemouche mouche des fruits mouche du vinaigre mouche noire moucheron
En rapport avec "motoneurone"

Un interneurone intègre les informations de chaque unité de couche d'entrée à laquelle il est connecté.

Le neurone désigne une cellule nerveuse spécialisée dans le traitement de l'influx nerveux, formé d'un corps cellulaire, de dendrites et d'un axone, de taille...

Un aleurone est un grain ovoïde, contenant des protides avec, dans certains cas, différentes inclusions; présent dans les cellules des graines mûres...