
La polyploïdie
La polyploïdie et l'autopolyploïdie sont une caractéristique d'un organisme possédant plus de deux jeux de chromosomes, contrairement à un organisme haploïde. Un organisme polyploïde peut être triploïde, tétraploïde ou hexaploïde.
Plusieurs cas de polyploïdie :
La ploïdie désigne le nombre de chromosomes présents dans un individu. La polyploïdie diffère d'une ploïdie simple des chromosomes haploïdes par le nombre augmenté de chromosomes (la tétraploïdie implique 4 chromosomes).
Généralités
La polyploïdie est fréquente chez les plantes et chez certains groupes de poissons et d'amphibiens. L'endoréplication de la chromatine libre en phase S (interphase) est toujours suivie d'une phase G1. L'endomitose avec polyténie débute par l'endoprophase, au cours de laquelle les chromosomes se condensent. Leurs chromatides filles se séparent et peuvent être comptées en endométaphase.
Les chromosomes polyploïdes (multiples) présentent un nombre de copies (4n, 8n, 16n, 32n, etc.) et possèdent une membrane nucléaire. Ces noyaux sont appelés tétraploïdes, octoploïdes, décaexiploïdes, triantadyoploïdes, etc. Selon ce nombre, la quantité totale d'ADN de ces noyaux cellulaires devrait être de 4C, 8C, 16C, 32C, etc.
Avantages
Étant donné la fréquence élevée de la polyploïdie chez certains taxons, comme les plantes, les poissons et les grenouilles, il est clair que la polyploïdie présente certains avantages. Chez les plantes, on observe fréquemment l'hétérosis (vigueur hybride), où la descendance polyploïde issue de deux progéniteurs diploïdes est plus vigoureuse et plus saine que chacun de ses parents diploïdes.
L'une d'elles est que l'appariement forcé des chromosomes homologues au sein d'un allotétraploïde empêche la recombinaison entre les génomes des progéniteurs initiaux, maintenant ainsi l'hétérozygotie au fil des générations. Cette hétérozygotie empêche l'accumulation de mutations récessives dans les génomes des générations suivantes, préservant ainsi la vigueur de l'hybride. Un autre facteur important est la redondance des gènes.
Schéma de la polyploïdie :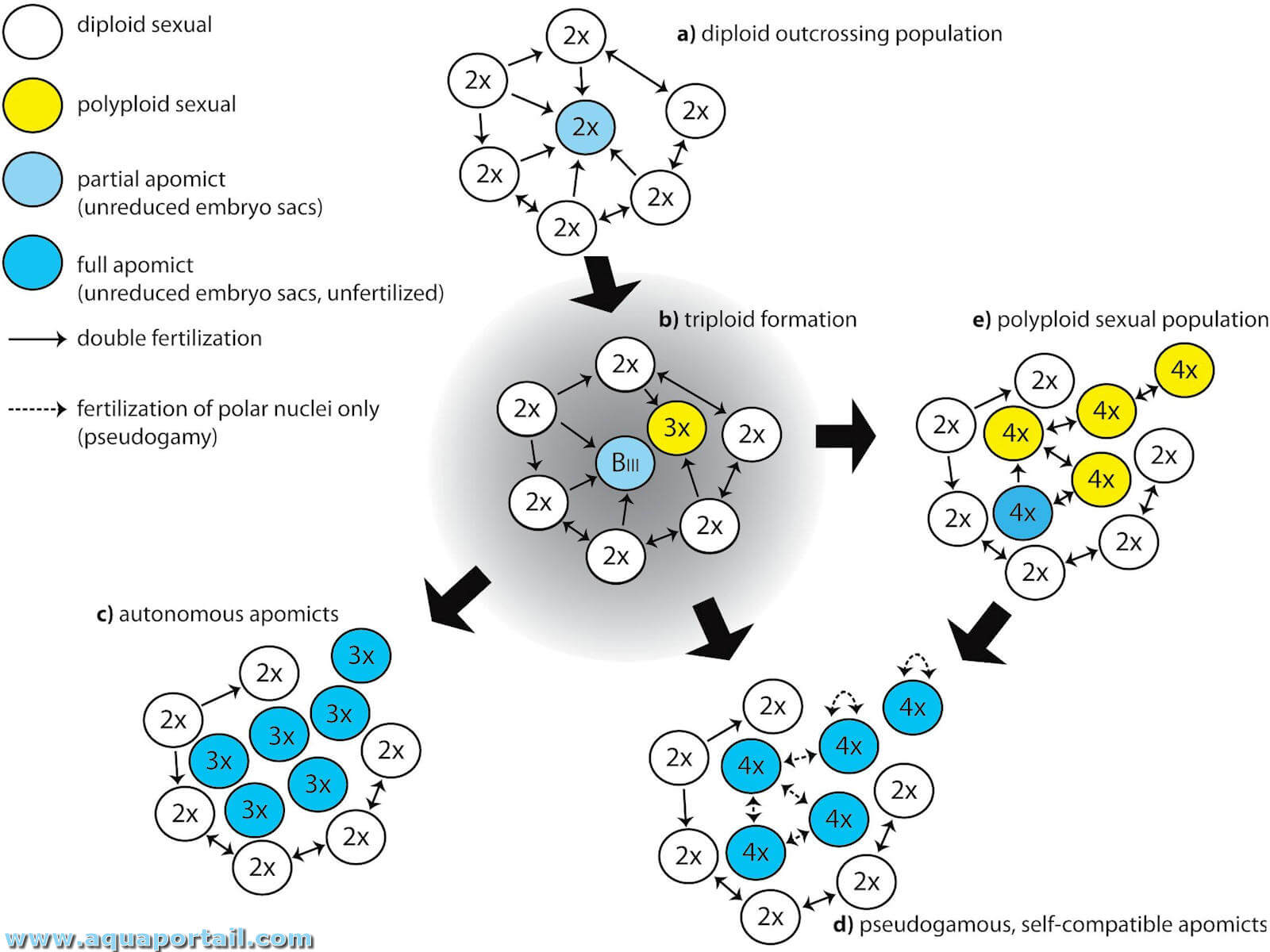
La polyploïdie génétique désigne un nombre de jeux de chromosomes supérieur à deux. Un organisme polyploïde peut être tétraploïde ou hexaploïde. Schéma des variantes de la polyploïdie.
Étant donné que la progéniture polyploïde possède maintenant deux fois plus de copies de tout gène particulier, la progéniture est protégée des effets néfastes des mutations récessives.
Ceci est particulièrement important au cours de la vie des gamétophytes. On pourrait imaginer que, durant la phase haploïde du cycle de vie, aucun allèle récessif d'une mutation délétère ne soit masqué par la présence d'un allèle dominant fonctionnant normalement, ce qui permettrait à la mutation de provoquer un arrêt du développement du pollen ou du sac d'oeufs. À l'inverse, un gamète diploïde permet de masquer cet allèle délétère par la présence de l'allèle normal dominant, protégeant ainsi le pollen ou le sac de l'oeuf d'un dysfonctionnement du développement. Cet effet protecteur de la polyploïdie pourrait être important lorsque de petites populations isolées sont forcées de se croiser.
Un autre avantage conféré par la redondance des gènes est la possibilité de diversifier la fonction des gènes au fil du temps. En d'autres termes, des copies supplémentaires de gènes qui ne sont pas nécessaires au fonctionnement normal de l'organisme pourraient finir par être utilisées de manières nouvelles et totalement différentes, ouvrant ainsi de nouvelles perspectives pour la sélection évolutive.
Fait intéressant, la polyploïdie peut affecter la sexualité de manière à offrir des avantages sélectifs. Une solution consiste à perturber certains systèmes d'auto-incompatibilité, permettant ainsi l'autofécondation. Cela pourrait être le résultat des interactions entre les génomes parentaux chez les allopolyploïdes. Une autre solution consiste à favoriser l'apparition de la reproduction asexuée, associée à la polyploïdie chez les plantes et les animaux. Ce changement de stratégie de reproduction peut améliorer la condition physique dans des environnements statiques.
Inconvénients
Malgré tous les avantages que la polyploïdie peut conférer à un organisme, il existe également un grand nombre d'inconvénients, observés et hypothétiques. L'un de ces inconvénients concerne les changements relatifs entre la taille du génome et le volume de la cellule. Le volume cellulaire est proportionnel à la quantité d'ADN dans le noyau cellulaire.
Par exemple, doubler le génome d'une cellule devrait doubler le volume d'espace occupé par les chromosomes dans le noyau, mais il ne provoque qu'une augmentation de 1,6 fois de la surface de l'enveloppe nucléaire. Cela peut perturber l'équilibre des facteurs qui interviennent normalement dans les interactions entre les chromosomes et les composants nucléaires, y compris les protéines liées à l'enveloppe. Le positionnement périphérique de l'hétérochromatine télomérique et centromérique peut également être perturbé, car il y a moins d'espace de surface relatif sur l'enveloppe nucléaire pour permettre ce positionnement.
La polyploïdie peut également être problématique pour l'achèvement normal de la mitose et de la méiose. D'une part, la polyploïdie augmente l'apparition d'irrégularités du fuseau, ce qui peut conduire à la ségrégation chaotique des chromatides et à la production de cellules aneuploïdes chez les animaux et les levures. Les cellules aneuploïdes, qui présentent un nombre anormal de chromosomes, sont plus facilement produites lors de maladies impliquant trois séries de chromosomes ou plus que dans les cellules diploïdes.
Les autopolyploïdes peuvent former de multiples arrangements de chromosomes homologues au niveau de la métaphase méiotique I, ce qui peut entraîner des schémas de ségrégation anormaux, tels que 3 : 1 ou 2 : 1 plus un retardataire. (Les chromosomes retardataires ne se fixent pas correctement à l'appareil fusilier et se séparent donc de manière aléatoire en cellules filles). Ces schémas de ségrégation anormaux ne peuvent pas être résolus en produits équilibrés, et la ségrégation aléatoire de plusieurs types de chromosomes produit principalement des gamètes aneuploïdes.
L'appariement des chromosomes lors de la méiose I est plus contraignant chez les allopolyploïdes que chez les autopolyploïdes, mais le maintien stable des deux compléments chromosomiques parentaux nécessite également la formation de gamètes équilibrés.
Un autre inconvénient de la polyploïdie comprend les modifications potentielles de l'expression des gènes. Il est généralement admis qu'une augmentation du nombre de copies de tous les chromosomes affecterait tous les gènes de manière égale et devrait entraîner une augmentation uniforme de l'expression des gènes. Les exceptions possibles incluent les gènes qui répondent à des facteurs régulateurs qui ne changent pas proportionnellement à la ploïdie. Nous avons maintenant des preuves expérimentales de telles exceptions dans plusieurs systèmes.
Dans un exemple intéressant, les chercheurs ont comparé les niveaux d'ARNm par génome pour 18 gènes de maïs 1X, 2X, 3X et 4X. Alors que l'expression de la plupart des gènes augmentait avec la ploïdie, certains gènes présentaient des écarts inattendus par rapport aux niveaux d'expression attendus. Par exemple, la saccharose synthase a montré l'expression proportionnelle attendue dans les tissus 2X et 4X, mais son expression était respectivement trois et six fois plus élevée dans les tissus 1X et 3X. Deux autres gènes ont montré des tendances similaires, bien que moins extrêmes. Au total, environ 10 % de ces gènes ont montré une sensibilité à la ploïdie impaire.
L'instabilité épigénétique peut constituer un autre défi pour les polyploïdes. L'épigénétique fait référence aux modifications du phénotype et de l'expression des gènes qui ne sont pas causées par des modifications de la séquence de l'ADN. Selon l'hypothèse du choc génomique, des perturbations du génome, telles que la polyploïdisation, peuvent entraîner des modifications généralisées de la régulation épigénétique. Bien qu'il existe peu d'instabilités épigénétiques documentées chez les autopolyploïdes, il existe quelques exemples intéressants qui méritent d'être mentionnés.
Dans un cas, la suppression des transgènes s'est produite plus fréquemment chez les tétraploïdes d'Arabidopsis thaliana que chez les diploïdes d'Arabidopsis thaliana, suggérant un effet de la ploïdie sur le remodelage des chromosomes. Cependant, plusieurs facteurs ne peuvent être exclus de l'observation de ce phénomène, notamment la duplication du puissant promoteur 35S du virus de la mosaïque du chou-fleur dans le transgène.
Dans un autre cas, l'activation d'un transposon d'ADN de la famille Spm/CACTA a été observée dans des autopolyploïdes. Malheureusement, la généralité de ce changement n'a pas pu être déterminée car plusieurs autopolyploïdes indépendants n'ont pas été examinés.
Inversement, de nombreuses preuves de remodelage épigénétique sont disponibles chez les allopolyploïdes. Des modifications génomiques structurelles, telles que la méthylation de l'ADN, et des modifications de l'expression accompagneraient la transition vers l'alloploïdie dans plusieurs systèmes végétaux, notamment Arabidopsis et le blé. Les informations les plus détaillées sont disponibles pour le système modèle Arabidopsis. Par exemple, dans un croisement de A. thaliana et A. arenosa, des gènes régulés épigénétiquement ont été identifiés en comparant les transcriptions des parents autotétraploïdes aux transcriptions de la descendance néoallopolyploïde. Les gènes d'A. thaliana affectés par la régulation épigénétique ont été définis comme ceux qui réagissaient au passage de l'autopolyploïdie à l'allopolyploïdie.
Au total, on estime que 2 à 2,5 % des gènes d'A. thaliana ont subi des modifications réglementaires au cours de la transition vers l'allopolyploïdie. Une étude plus détaillée sur des micropuces a examiné la régulation de 26 000 gènes chez des néoallopolyploïdes d'Arabidopsis a détecté une divergence du transcriptome entre les progéniteurs de plus de 15 %, due à des gènes fortement exprimés dans Arabidopsis thaliana et non pas dans Arabidopsis arenosa.
De manière significative, l'expression d'environ 5 % des gènes a divergé de la valeur du milieu parent dans deux allotétraploïdes dérivés indépendamment, ce qui est cohérent avec la régulation génique non additive après l'hybridation (Wang et al., 2006). Pris ensemble, ces résultats suggèrent que le syndrome d'instabilité des néoallopolyploïdes peut être attribué principalement à une divergence de régulation entre les espèces parentales, conduisant à des incompatibilités génomiques chez les descendants allopolyploïdes.
L'aneuploïdie pourrait également jouer un rôle dans le remodelage épigénétique des néoallopolyploïdes, soit en modifiant le dosage de facteurs codés par des chromosomes dont le nombre de copies est supérieur ou inférieur au nombre attendu, ce qui entraîne des modifications des loci imprimés, ou en exposant des régions de chromatine non appariées à des régions épigénétiques. mécanismes de remodelage.
Dans le dernier cas, cette susceptibilité de l'ADN méiotiquement non apparié au silence a été signalée pour la première fois pour le champignon Neurospora crassa, mais il semble s'agir d'un phénomène général. Une partie de l'instabilité épigénétique observée chez les allopolyploïdes pourrait donc résulter de l'aneuploïdie.
Conclusions
Des études récentes ont fourni des informations intéressantes sur les conséquences réglementaires et génomiques de la polyploïdie. Parallèlement aux preuves émergentes d'une duplication ancestrale par la polyploïdisation chez des plantes modèles, des champignons et des espèces animales, la connaissance de ces conséquences a stimulé la réflexion sur la relation entre les premiers événements de polyploïdisation, le succès de la polyploïdie et le devenir à long terme de nouvelles espèces.
En rapport avec "polyploïdie"
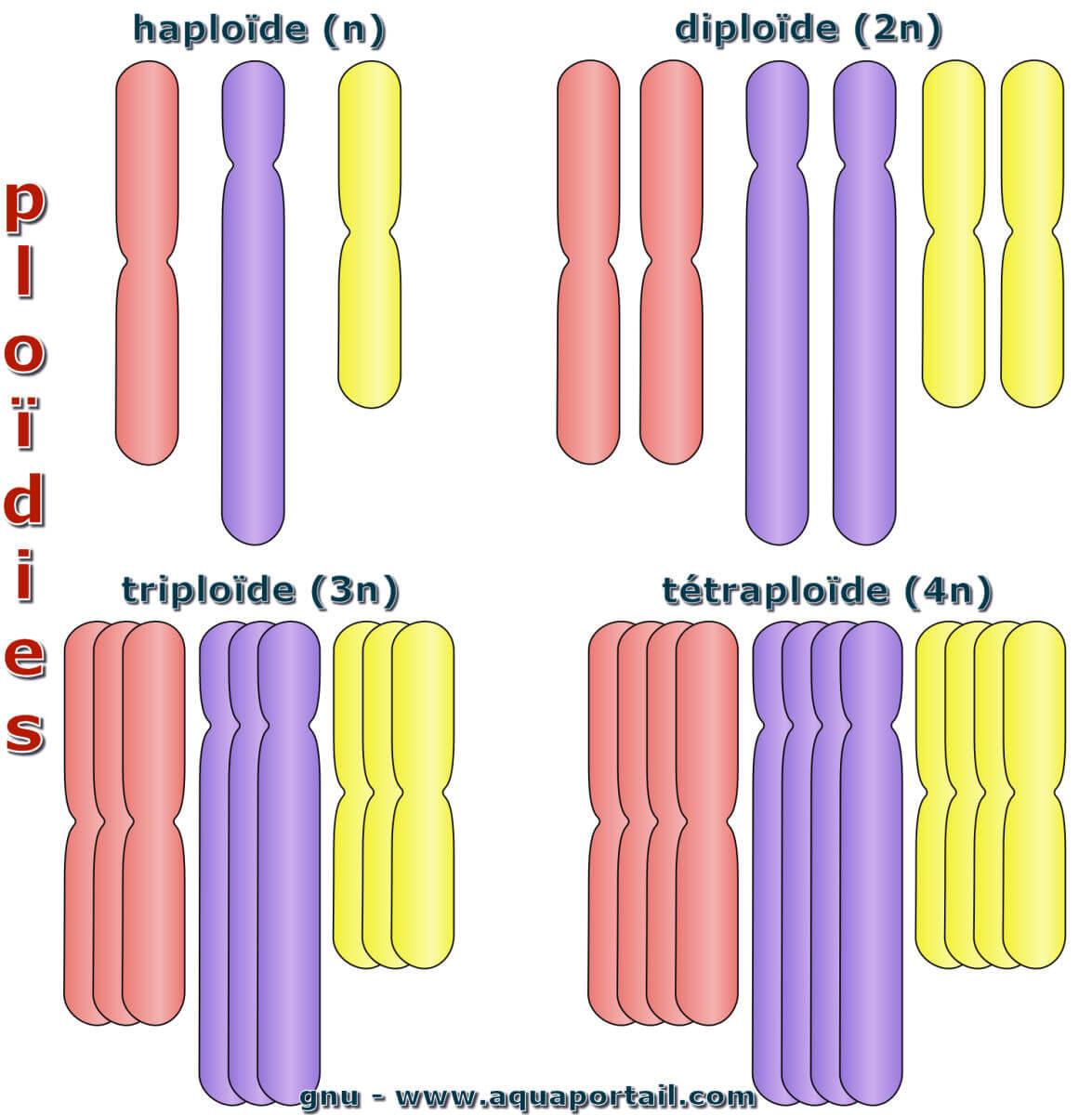
Un organisme haploïde a des cellules possèdant 1n chromosome (nombre minimal de base). L'haploïde possède un ensemble distinct de chromosomes non appariés...
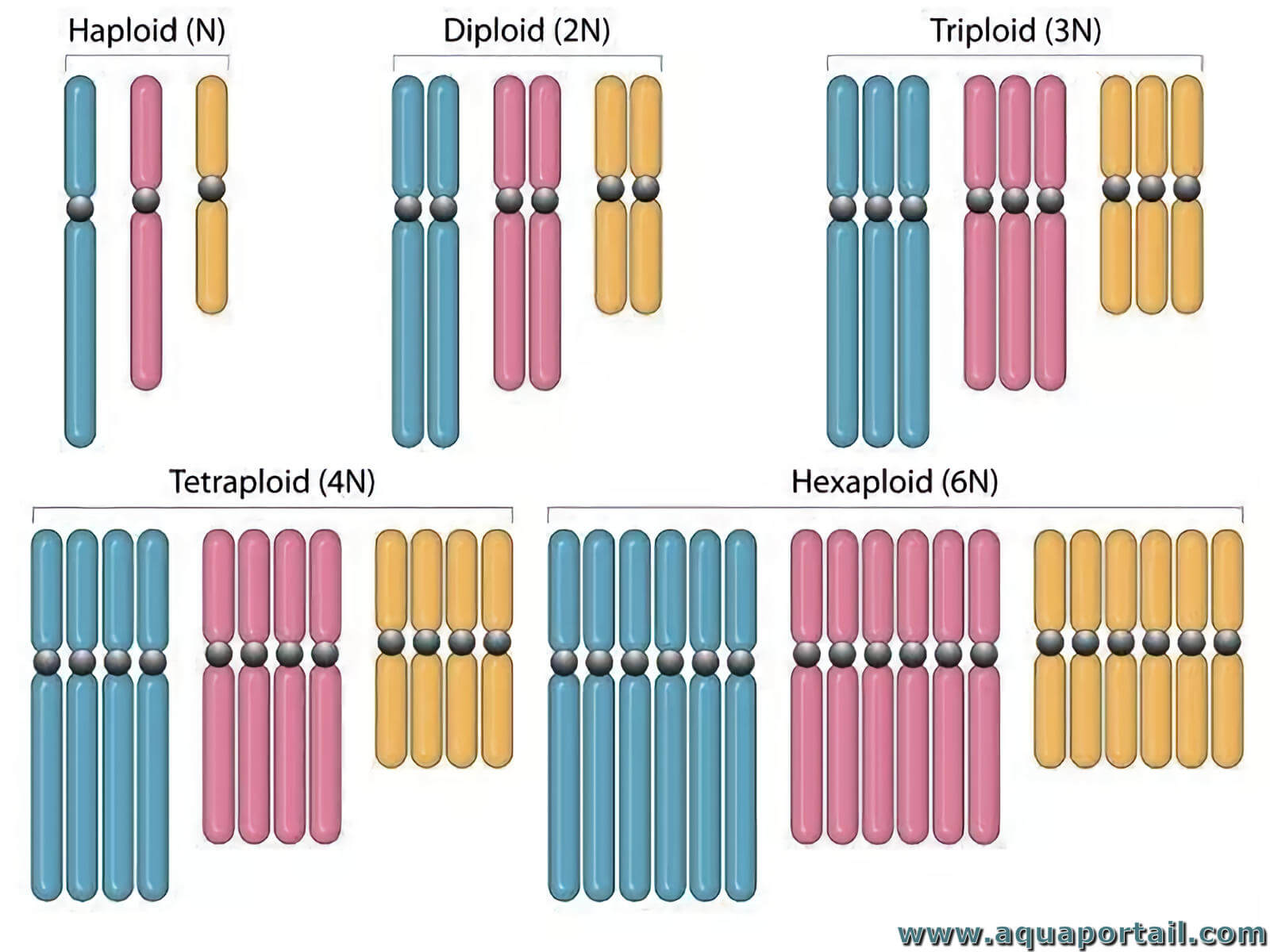
Un polyploïde est un organisme dont les cellules possèdent un multiple de n chromosomes. La polyploïdie est le caractère d'une cellule possédant plus...
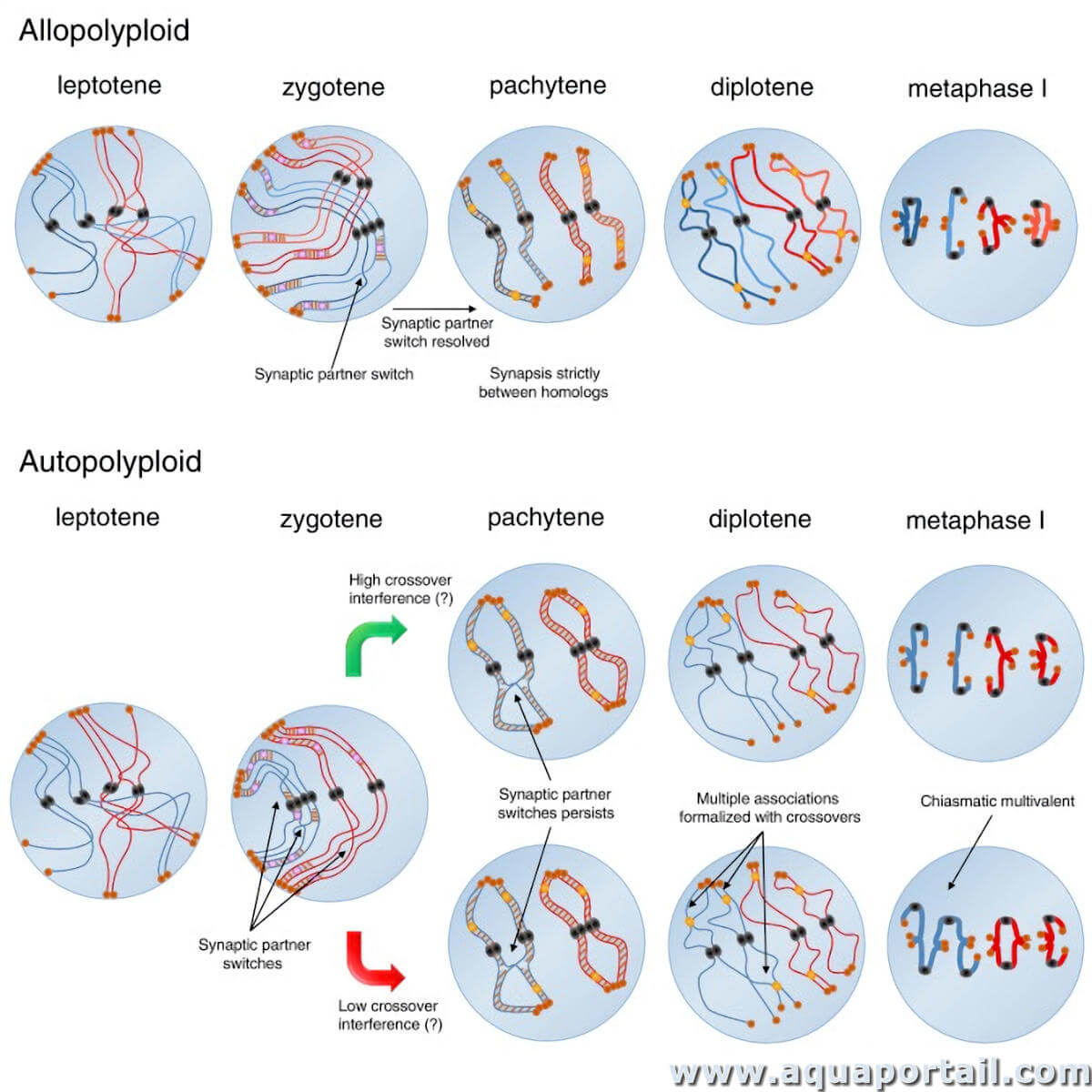
Un organisme allopolyploïde qualifie un organisme dont le noyau de chacune des cellules somatiques renferme un nombre de chromosomes supérieur à 2n.

L'amphidiploïdie est en rapport avec un double diploïde lors d'une division cellulaire en méiose, notamment chez les hybrides : voir un allopolyploïde.