
Évolution et histoire des feuilles avec la photosynthèse
Les feuilles ont évolué avec la photosynthèse depuis 380 millions d'années:Les feuilles, en tant que site de la photosynthèse, sont essentielles à la croissance et à la survie des plantes. Il y a environ 380 millions d'années, les plantes dotées de tissus vasculaires (trachéophytes) ont d'abord développé un type spécial de feuille, appelé microphylle. La photosynthèse foliaire débutait avec les microphylles, et évoluait ensuite avec les mégaphylles. Ces évolutions intègrent la théorie du télome.
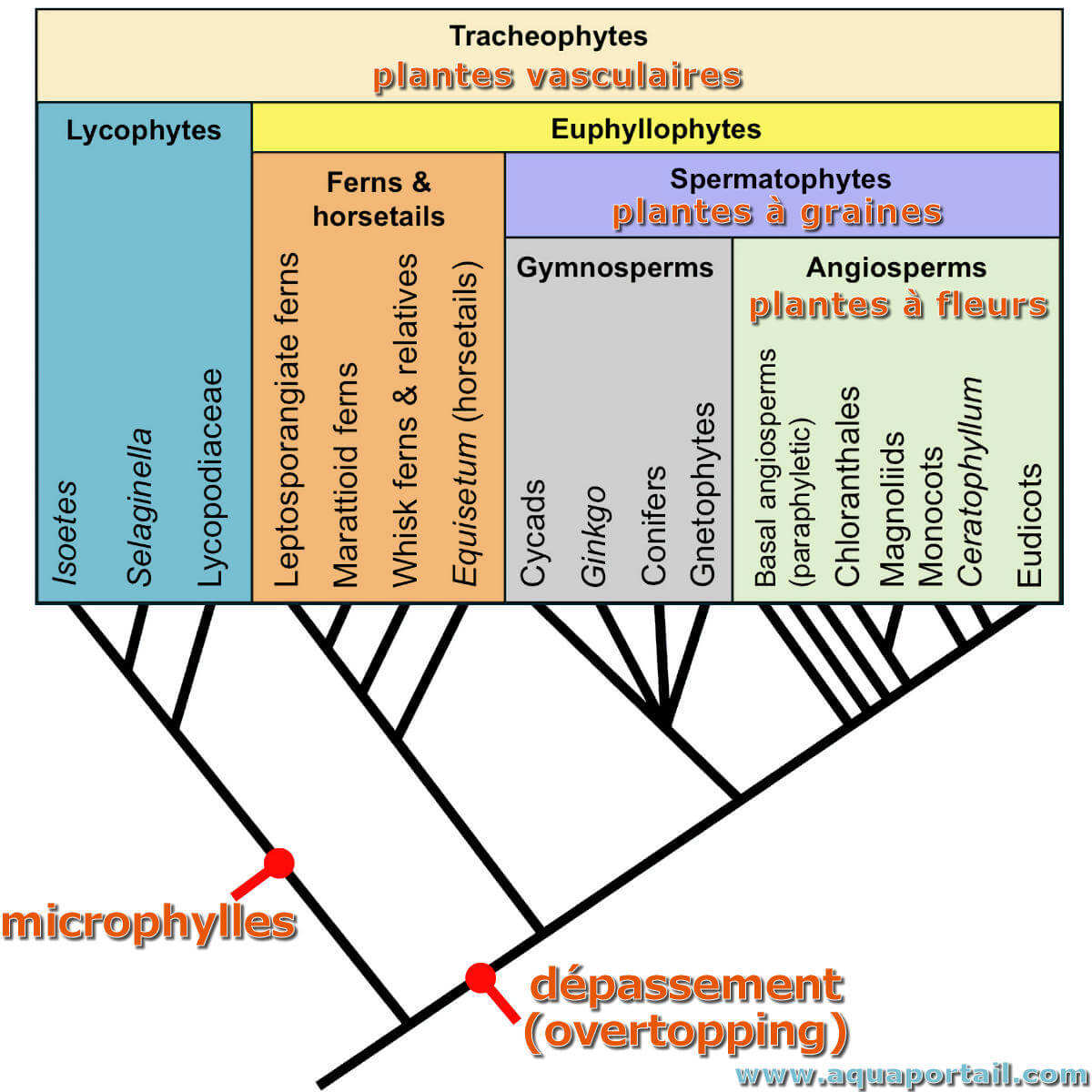
L'arbre des relations entre les trachéophytes vivants (plante vasculaire) ci-dessus montre les étapes clés de l'origine des feuilles avec l'évolution photosynthétique.
La variation évolutive va des séquences de développement au sein d'une pousse à la réponse au microenvironnement au sein de la couronne, de la variation entre les espèces au sein et entre les communautés, à la variation à travers les lignées évolutives.
Les plantes vasculaires évoluent. Les plantes ont développé un certain nombre d'adaptations qui les ont aidées à faire face à ces problèmes sur la Terre ferme. L'une des premières et des plus importantes a été l'évolution des tissus vasculaires. Les tissus vasculaires forment le "système de plomberie" d'une plante. Ils transportent l'eau et les minéraux du sol vers les feuilles pour la photosynthèse.
Deux types de feuilles de base sont présents dans les plantes vasculaires, les microphylles (petite feuille) et les mégaphylles (grande feuille). Ces termes sont utilisés pour distinguer les types de feuilles qui diffèrent par leurs origines évolutives et, souvent, leurs attributs structurels. Certains scientifiques ont critiqué ces termes comme étant déroutants, car les petits mégaphylles simples ressemblent aux microphylles. De plus, les termes sont parfois utilisés de manière purement descriptive sans implications évolutives. Ainsi, il a été proposé que les deux principaux types de feuilles des plantes vasculaires soient plutôt appelés lycophylles (= microphylles) et euphylles (= mégaphylles).
Un microphylle a généralement une seule nervure médiane et provient d'une tige qui n'a pas d'espaces foliaires, dans les régions du tissu parenchymateux (c'est-à-dire non spécialisé) où le brin vasculaire mène à la base de la feuille. Le microphylle peut provenir d'une excroissance d'une tige vascularisée ou d'une simplification évolutive d'un système de branches complexe. Les feuilles de certaines plantes modernes des Lycopodophytes (Lycopodes) et des Sphénophytes (prêles) sont classées comme microphylles. Bien que les microphylles de ces plantes modernes soient assez petits, certains de leurs parents fossilisés avaient de très gros microphylles.
30 millions d'années plus tard, soit il y a environ 350 millions d'années, les plantes ont d'abord développé des mégaphylles, le type de feuille des plantes à graines et des fougères modernes. Les microphylles sont une synapomorphie pour les Lycophytes. Le dépassement, l'une des étapes de l'origine hypothétique des mégaphylles, est une synapomorphie pour les euphyllophytes.
Le dépassement (overtopping en anglais) est l'une des premières étapes de l'évolution hypothétique du mégaphylle, est considéré comme une synapomorphie pour les euphyllophytes.
Un mégaphylle a généralement un motif de nervation complexe et provient d'une tige qui présente des espaces foliaires ou des régions de tissu de parenchyme où le brin vasculaire mène à la base de la feuille. Une théorie propose que les mégaphylles, ainsi que d'autres organes végétaux, ont évolué par modification des systèmes de branches. En d'autres termes, les mégaphylles peuvent avoir évolué par aplatissement d'un système de branches tridimensionnel et connexion des branches aplaties avec une sangle cellulaire.
L'évolution des premières avec la plante Cooksonia :
Illustration de Cooksonia, la plus ancienne plante vasculaire connue du Silurien moyen. L'encart est un dessin au trait illustrant l'habitude de ramification dichotomique typique (A), une coupe transversale de l'axe montrant le tissu vasculaire (B) et un détail des télomes terminaux, chacun avec un sporange (C). L'arrière-plan est le rendu d'un artiste d'un paysage dominé par Cooksonia.
De nombreux botanistes pensent que les quatre verticilles différents d'une fleur (sépales, pétales, étamines et carpelles) proviennent d'une modification évolutive des mégaphylles d'une plante à spores libres. Peu de temps après l'origine évolutive des Angiospermes (plantes à fleurs), il y a eu une division majeure entre les monocotylédones (plantes dont les embryons ont un cotylédon) et les dicotylédones (plantes dont les embryons ont deux cotylédons). Les feuilles des monocotylédones modernes, comme les graminées et les lys, ont tendance à être étroites et à nervures parallèles; les feuilles des dicotylédones ont tendance à être larges et à nervures réticulées.
De nombreuses plantes modernes ont développé des feuilles complexes et hautement spécialisées. Par exemple, les organes insectivores des plantes carnivores sont tous des feuilles hautement spécialisées. Dischidia rafflesiana, une épiphyte tropicale, possède l'une des feuilles les plus spécialisées de toutes les plantes. Ses feuilles sont de forme tubulaire et elles recueillent les débris forestiers et l'eau de pluie, offrant un habitat aux colonies de fourmis qui vivent à l'intérieur. Au fur et à mesure que les fourmis meurent, leurs corps se dissolvent et les racines spéciales de la plante absorbent les nutriments qui sont libérés, fournissant ainsi de la nourriture à la plante.
La théorie du télome a été formulée par Walter Zimmermann dans les années 1930 et développée au cours des 30 années suivantes avec une bonne explication générale pour le public anglophone par Wilson (1953). Fondamentalement, la théorie tente d'expliquer les origines de tous les principaux organes végétaux comme des dérivations des structures simples des premières plantes terrestres du Silurien supérieur et du Dévonien inférieur. Ces plantes étaient déjà passées du gamétophyte photosynthétique bryophyte à un sporophyte photosynthétique et persistant. Ces petites plantes dressées n'étaient rien de plus que des axes à ramifications dichotomiques (tiges et racines), dont certains se terminaient par des sporanges. Zimmermann a qualifié les axes végétatifs intermédiaires de mésomes et les axes terminaux qu'il a appelés télomes.



