
Chromatine
Définition
La chromatine est un complexe d'ADN, d'ARN et de protéines qui forme les chromosomes dans le noyau des cellules eucaryotes. Elle constitue l'état physiologique du génome. Il en existe deux types : l'euchromatine décondensée et l'hétérochromatine condensée.
La chromatine :
La chromatine est un mélange d'ADN et de protéines qui forment les chromosomes présents dans les cellules des organismes supérieurs.
Explications
La chromatine est une association de l'ADN, de l'ARN et des protéines présentes dans le noyau interphase des cellules eucaryotes. En fonction de la manière dont l'ADN est présenté dans un noyau de cellule, elle est appelée euchromatine, décondensée et peu colorable, ou hétérochromatine, condensée et très colorable. Seule l'euchromatine est transcrite. L'unité structurelle de base de la chromatine est le nucléosome.
La structure de la chromatine est importante pour plusieurs processus différents tels que la stabilité du génome, l'expression des gènes, la réplication du génome et la réparation et la recombinaison de l'ADN.
La chromatine désigne un complexe constitué par l'ADN et les protéines chromosomiques (histones et protéines non-histones). Elle est donc de l'ADN filiforme non concentré, présente dans les cellules eucaryotes en interphase. Chez les cellules procaryotes, le noyau cellulaire contient de l'ADN entassé.
Dans une chromatine, la molécule d'ADN est associée à diverses protéines dont les histones qui sont organisées en nucléosomes. La molécule d'ADN s'enroule autour de chaque nucléosome en réalisant deux tours complets et se prolonge jusqu'au nucléosome suivant par un ADN internucléosomique.
Ne pas confondre chromatine et chromatide !
La structure fondamentale de la chromatine est essentiellement la même chez presque tous les eucaryotes, des levures unicellulaires aux organismes multicellulaires les plus complexes (à l'exception de la chromatine des dinoflagellés et des spermatozoïdes de vertébrés). Son rôle fondamental est de compacter le génome sous une forme suffisamment compacte pour permettre à des molécules d'ADN relativement grandes de tenir dans le noyau de la cellule.
lire plus de détails pour la chromatine...
Synonymes, antonymes
Voir tous les synonymes pour "chromatine".3 synonymes (sens proche) de "chromatine" :
0 antonyme (sens contraire).
Les mots ou les expressions apparentés à CHROMATINE sont des termes qui sont directement liés les uns aux autres par leur signification, générale ou spécifique.
Le mot CHROMATINE est dans la page 5 des mots en C du lexique du dictionnaire.
Mots en C à proximité
choroïde chorologie chott chromatide chromatides soeurs chromatinechromatographie chromatographie en phase gazeuse chromatophore chromatosome chromogène
En rapport avec "chromatine"
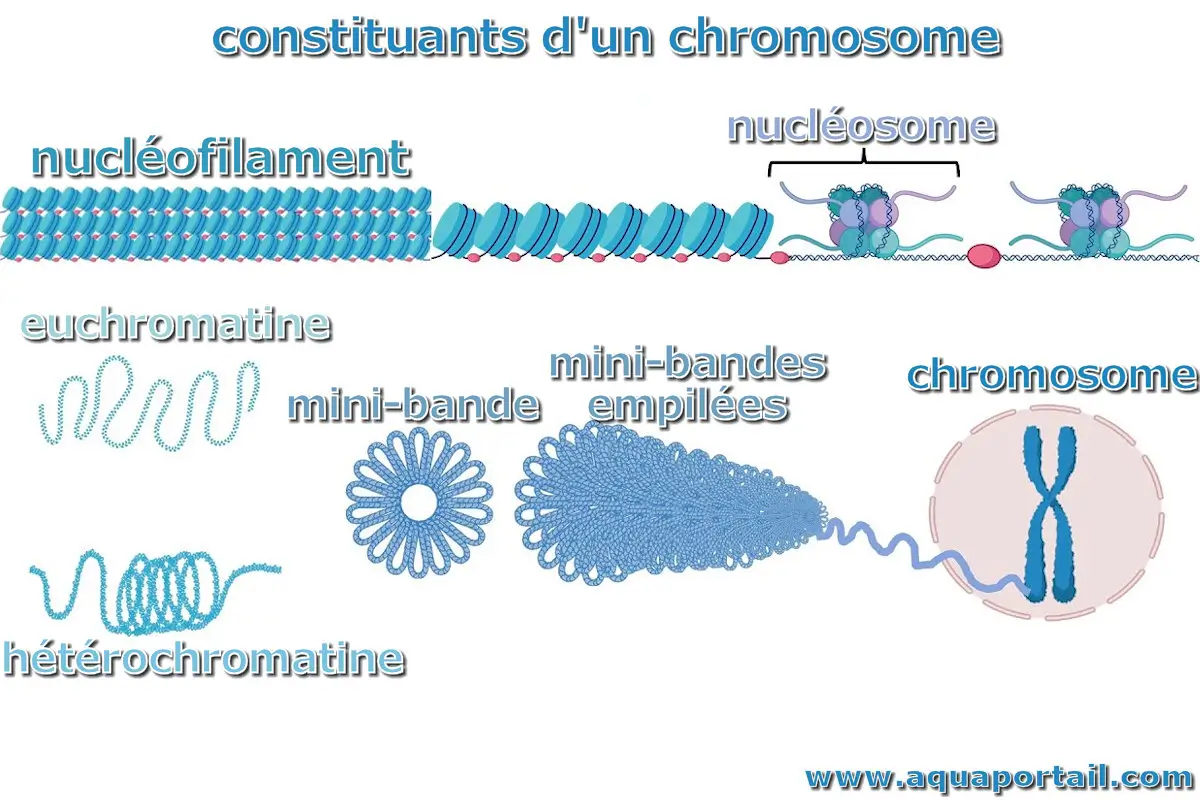
Un nucléofilament est le constituant de forme filamenteuse de la chromatine des chromosomes.

Une chromatide est chacun des deux brins de nucléofilament porteur d'une double hélice d'ADN lorsqu'un chromosome se divise longitudinalement en mitose...

Les chromatides soeurs sont deux copies générées du même chromosome lorsqu'il est répliqué pendant la phase de synthèse de l'interphase, avant le début...

La condensation de la chromatine correspond à la spiralisation des filaments de chromatine, aboutissant à des enroulements très serrés qui sont...